Immunohistochemical Study of the Distribution of Transcriptional Factor Sox2 in the Lingual Epithelium of Human Embryos and Fetuses
Transcriptional factor Sox2 is one of the key factors in the development of mammal sensory system. Sox2 expression was revealed in rodent developing taste buds, while the role of this factor in morphogenesis of human taste system still remains unstudied.
The aim of investigation was to study the character of a transcriptional factor Sox2 distribution in the lingual epithelium of human embryos and fetuses.
Materials and Methods. We carried out an immunohistochemical study of lingual epithelium of 28 human fetuses and embryos from 6th to 21st week of prenatal development.
Results. Immunopositive reaction to Sox2 was revealed in lingual epithelial cells starting from the 10th week of development and in all later stages. The greatest number of Sox2 immunopositive nuclei was localized in a basal layer of lingual epithelium and in epithelial evaginations associated with papillae morphogenesis. Since the 15th week of prenatal development Sox2 expression level increased in apical parts of taste papillae and in taste bud cells. The comparison of immunopositive nuclei distribution with the main morphogenetic events in lingual epithelium of human fetus showed Sox2 to regulate the morphogenesis of human lingual papillae and at early stages (before the 10th week) and not participate in differentiation of taste bud cells.
Mammalian taste buds are the clusters of receptor cells transforming chemical stimuli into a nerve impulse. Taste receptor cells like neurons are able to generate action potentials, form synapses with nerve endings and excrete neurotransmitters into a synaptic cleft [1, 2]. However, unlike neurons, taste bud cells are the descendants of the same cell line as epithelial cells surrounding a taste bud [3]. As well as other epithelial cells they are able to regenerate and renew themselves [4]. Due to such a unique combination of taste buds properties the investigation of their early embryonal differentiation is of interest for understanding morphogenesis mechanisms of both mammalian epithelial and sensor organs.
Early differentiation of lingual epithelium currently is most completely studied in rodents. The formation of taste organs in mice begins in the middle of intrauterine development when epithelial enlargements appear on the dorsal lingual surface called placodes [5]. With the development period increase the placodes evaginate and form taste papillae in the epithelium of which taste buds appear by the end of intrauterine development [5, 6]. Recently, the development of epithelial placodes and rodent taste papillae was shown to be accompanied by the expression of epithelial and mesenchymal signal molecules [2, 7]. However the question of a role of epithelial-mesenchymal regulation in taste bud morphogenesis initiation during embryonal development of mammalians remains controversial. Some authors consider the contact of nerve fibers with lingual epithelium to be necessary for the beginning of taste bud cells differentiation [8, 9], while the others believe that taste bud progenitor cells are already determined on early stages of taste placodes development under control of focal signals [10–12]. Over the last 10 years in lingual epithelium of mice embryos there have been found numerous factors and signal molecules participating in regulation of processes of taste buds and papillae differentiation, Sox2 being one of them [2, 6, 13]. Sox2 or SoxB1-HMG-box transcription factor plays an important role in mammalian development. It is abundantly expressed in the anlage of mammalian central nervous system and is also necessary for the development of retinal and inner ear sensory cells [14–16]. High level of Sox2 expression in mice lingual epithelium is found in epithelial placode cells, in fungiform papillae epithelium and in mature taste buds [6]. Hypomorphic mutations in mice have shown that if the level of Sox2 expression reaches only 20% from the norm the epithelial placodes form but mature taste papillae and buds are missing [6]. Represented facts suggest that Sox2 is one of the key factors in taste papillae morphogenesis regulation. However, it is difficult to assess the participation of Sox2 factor in differentiation initiation of mice taste bud cells due to chronologic convergence of processes of taste buds and papillae morphogenesis in rodents.
The early intrauterine development of human lingual taste buds and papillae differs from that in rodents. Differentiation of human lingual epithelium starts from the penetration of sensor nerve endings into the epithelium, and formation of clusters of elongated cells called taste bud primordia [9]. The difference of taste primordia from the epithelial placodes observed in the development of taste papillae in rodents is in the formation of synapses between primordial cells and nerve endings penetrated into epithelium that enables to consider primordia to be immature taste buds developing before taste papillae formation [8]. Since morphogenesis processes of taste buds and papillae at early human developmental stages are separated in time, a human tongue is a convenient object to test lingual epithelial cell differentiation models described in rodents.
The aim of investigation was to study the character of a transcriptional factor Sox2 distribution in the lingual epithelium of human embryos and fetuses.
Materials and Methods. The work was performed on abortive material of 28 human embryos and fetuses from the 6th to 21st week of intrauterine development from the collection of nerve system development of RS of Human Morphology of RAMS. The material was fixed in a 4% paraformaldehyde solution. Gestational age was determined by medical documents, coccygeal-temporal size, body weight, rostrocaudal length and descriptive signs in accordance with recognized techniques [17].
At early developmental stages (before the 13th week) the material of a whole head was used in investigation, at later stages the oral cavity with palatinal and mandibular bones were separated. If appropriate the material was placed into a decalcinated mixture based on azotic acid. The material was embedded into paraffin, and serial slices 10 µm thick were prepared. Subsequently, the slices were studied immunohistochemically with primary antibody to Sox2 (Anti-Sox2 antibody, polyrabbit, Abcam, USA). In immunohistochemical treatment we used UltraVision LP Detection System HRP Polymer (LabVision, Great Britain). In the process of immunohistochemical study, dewaxed, hydrated slices were treated by 3% H2O2 within 20 min for endogen peroxidase blocking. Further, the slices were heat treated in 0.01 M citrate buffer (pH=6.0) within 5 min for antigen disclosure. The slices were processed by UltraBlock from the kit used for 5 min for unspecific binding blocking. The slices were incubated with the primary antibody in a moist chamber at 37°C within 1 h. Further, the reagents from the kit were used according to manufacturer specification. Reactions with the substitution of primary antibody 0.01 M by the phosphate-saline buffer solution — PBC (pH=7.3–7.5) served as negative control.
Results. Primary taste primordia develop in human embryo lingual epithelium in the beginning of the 7th week of intrauterine development. At early gestational age (from the 6th to the 9th week) clear immunepositive reaction to Sox2 factor in lingual dorsal epithelium is absent. Starting from the 8th week of intrauterine development mild immunopositive reaction presents in human lingual epithelial cells (See Fig., a), but since the marker was found not only in nuclei but in cell cytoplasm the epithelium stain at early developmental stages may result from nonspecific cross reaction that requires further studies.
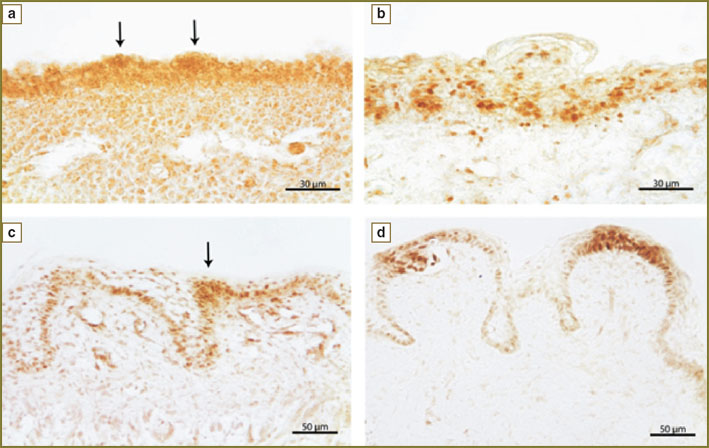
|
The human lingual epithelium on week 8 (a), 10 (b), 14 (c) and 18 (d) of intrauterine development, reaction with antibodies to Sox2 transcription factor. Arrows indicate taste primordia (a) and taste buds (c) |
Since the 10th week of intrauterine development clear and contrast reaction to Sox2 is revealed in the dorsal epithelium (See Fig., b). According visual assessment most stained nuclei are seen in the basal epithelial layer. Solitary spread immunopositive to Sox2 cells are found in superior epithelial layers. The number of such cells increases in places where an epithelial layer bends due to the formation and growth of lingual papillae. No differences in this marker expression depending on papillae type were found at this development stage.
On the 12–14th week of intrauterine development the distribution of immunopositive to Sox2 cells in epithelium becomes more disperse (See Fig., c). In the lingual basal epithelial layer their number decreases, and in more superficial layers still there are stained nuclei but they are not so numerous compared to the previous developmental stages. A great number of immunopositive nuclei persist in the basal epithelium of developing lingual papillae. Immunopositve reaction to Sox2 is also found in nuclei of taste bud cells.
By the 15th week of intrauterine development the intensity of reaction to Sox2 in the basal layer of lingual epithelium starts reducing. Against a mild immunopositive reaction, in lingual epithelium there stand out the apical parts of papillae, on which taste buds are located. The taste bud cell nuclei, as well as those of epithelial cells surrounding them are brightly stained. Iimmunoreactivity to Sox2 also remains in the basal parts of lingual papillae.
From the 18th till the 21st week of intrauterine development a bright and clear reaction to Sox2 is observed only on the top of taste papillae (See Fig., d). Immunopositive to Sox2 nuclei are situated in taste bud cells and in epithelial cells surrounding them. As far as taste buds become distant the reaction in epithelial cells of the apical part of papillae decreases.
Discussion. Primordial taste buds develop in the beginning of the 7th week of human intrauterine development that is consistent with the findings of other authors [8]. At the stage of taste primordia (before the 10th developmental week) no clear immunopositive reaction to Sox2 in the lingual epithelium was found that may result from low expression of this marker. Following the 10th week of intrauterine development there starts active morphogenesis of lingual papillae and there is a sharp rise of Sox2 expression. Most nuclei immunopositive to Sox2 are localized in basal epithelial layer, as well as in the growing basal parts of papillae, i.e. in places of active proliferation and differentiation of epithelial cells. The obtained results are in agreement with data of experiments carried out on mice. Mutant mice with Sox2 overexpression had increased number of fungiform papillae rather than taste buds [6]. At early developmental stages Sox2 appears to regulate the morphogenesis of lingual papillae and takes no part in initiation of taste bud cell differentiation. Starting from the 15th week of human intrauterine development intense Sox2 expression is found in the epithelium of apical parts of fungiform papillae and in taste bud cells. Such distribution of the marker is consistent with the results of investigations performed on mature mice, in which Sox2 is expressed only in taste bud cells and epithelial cells surrounding them [18]. Sox2 is likely to participate in the processes of maturation, renewal and regeneration of mammalian taste buds. The dependency of Sox2 expression on innervation is proved by experiments on mature mice, in which expression of this factor discontinues in degradation of gustative nerves [18]. At early developmental stages prior to penetration of nerve endings into mice lingual epithelium, Sox2 expression in placodes of lingual papillae is controlled by Wnt signal pathway [2]. Wnt activation is one of the unstudied problems since in oral epithelium at the same stages of embryonal development it results in different consequences such as teeth or lingual papillae formation [2]. Most studies in this field have been performed on rodents, and the results of few works on human embryos indicate the interspecies differences in expression of factors and signal molecules. For example, Sox2 expression is found in the development of germ cells in mice embryonal gonads, while there is no Sox2 expression in human embryos [19]. All abovementioned suggests interspecies interpolations to be insufficient to understand human morphogenesis processes, and further studies on transcription factors and signal systems on the human embryonal material are required.
Conclusion. Sox2 transcription factor is found in the lingual epithelium of all studied human embryos starting from the 10th week of intrauterine development. The comparison of distribution of immunopositive nuclei with the main morphogenetic events in the human embryos lingual epithelium showed Sox2 to regulate the morphogenesis of lingual papillae and take no part in differentiation of taste bud cells at early developmental stages (before the 10th week).
Study Funding. The work was supported by “Fundamental” Fund, a specialized Capital Management Fund for Research Support in the Sphere of Biology and Medicine.
Conflict of Interests. The author declares no conflict of interest.
References
- Finger T.E., Danilova V., Barrows J., Bartel D.L., Vigers A.J., Stone L., Hellekant G., Kinnamon S.C. ATP signaling is crucial for communication from taste buds to gustatory nerves. Science 2005; 310: 1495–1499.
- Lui F., Millar S.E. Wnt/β-catenin signaling in oral tissue development and disease. J Dent Res 2010; 89(4): 318–330, http://dx.doi.org/10.1177/0022034510363373.
- Hirota M., Ito T., Okudela K., Kawabe R., Hayashi H., Yazawa T., Fujita K., Kitamura H. Expression of cyclin-dependent kinase inhibitors in taste buds of mouse and hamster. Tissue Cell 2001; 33: 25–32.
- Hamamichi R., Asano-Miyoshi M., Emori Y. Taste buds contains both short-lived and long-lived cell populations. Neuroscience 2006; 141: 2129–2138.
- Mbiene J.P. Taste placodes are primary targets of geniculate but not trigeminal sensory axons in mouse developing tongue. J Neurocytol 2004; 33: 617–629.
- Okubo T., Pevny L.H., Hogan B.L.M. Sox2 is required for development of taste bud sensory cells. Genes & Development 2006; 20: 2654–2659, http://dx.doi.org/10.1101/gad.1457106.
- Hall J.M., Bell M.L., Finger T.E. Disruption of sonic hedgehog signaling alters growth and patterning of lingual taste papillae. Dev Biol 2003; 255: 263–277, http://dx.doi.org/10.1016/S0012-1606(02)00048-9.
- Oakley B., Witt M. Building sensory receptors on the tongue. J Neurocytol 2004; 33(6): 631–646, http://dx.doi.org/10.1007/s11068-005-3332-0.
- Tamatsu Y., Gasser R.F. Development of the sensory nerves to the dorsum of the tongue in staged human embryos. Clinical Anatomy 2004; 17: 99–106.
- Mbiene J.P., Roberts J.D. Distribution of keratin 8-containing cell clusters in mouse embryonic tongue: evidence for a prepattern for taste bud development. J Comp Neurol 2003; 457(2): 111–122.
- Zhou Y., Liu H.X., Mistretta C.M. Bone morphogenetic proteins and noggin: inhibiting and inducing fungiform taste papilla development. Dev Biol 2006; 297(1): 198–213, http://dx.doi.org/10.1016/j.ydbio.2006.05.022.
- Thirumangalathu S., Barlow L.A. In vivo fate tracing studies of mammalian. Taste cell progenitors international symposium on olfaction and taste. Ann NY Acad Sci 2009; 1170: 34–38, http://dx.doi.org/10.1111/j.1749-6632.2009.04371.x.
- Liu F., Thirumangalathu S., Gallant N.M., Yang S.H., Stoick-Cooper C.L., Reddy S.T., Andl T., Taketo M.M., Dlugosz A.A., Moon R.T., Barlow L.A., Millar S.E. Wnt-beta-catenin signaling initiates taste papilla development. Nature genetics 2007; 39(1): 106–112, http://dx.doi.org/10.1038/ng1932.
- Van Raay T.J., Moore K.B., Iordanova I., Steele M., Jamrich M., Harris W.A., Vetter M.L. Frizzled 5 signaling governs the neural potential of progenitors in the developing Xenopus retina. Neuron 2005; 46(1): 23–36, http://dx.doi.org/10.1016/j.neuron.2005.02.023.
- Taranova O.V., Magness S.T., Fagan B.M., Wu Y., Surzenko N., Hutton S.R., Pevny L.H. Sox2 is a dose-dependent regulator of retinal neural progenitor competence. Genes & Development 2006; 20: 1187–1202, http://dx.doi.org/10.1101/gad.1407906.
- Kiernan A.E., Pelling A.L., Leung K.K., Tang A.S., Bell D.M., Tease C., Lovell-Badge R., Steel K.P., Cheah K.S. Sox2 is required for sensory organ development in the mammalian inner ear. Nature 2005; 434(7036): 1031–1035, http://dx.doi.org/10.1038/nature03487.
- Savel’ev S.V. Stadii embrional’nogo razvitiya mozga cheloveka [The stages of human brain embryogenesis]. Moscow: VEDI, 2002; 112 p.
- Suzuki Y. Expression of Sox2 in mouse taste buds and its relation to innervation. Cell Tissue Res 2008; 332: 393–401, http://dx.doi.org/10.1007/s00441-008-0600-1.
- Perrett R.M., Turnpenny L., Eckert J.J., O’Shea M., Sonne S.B., Cameron I.T., Wilson D.I., Rajpert-De Meyts E., Hanley N.A. The early human germ cell lineage does not express SOX2 during in vivo development or upon in vitro culture. Biol Reprod 2008 May; 78(5): 852–858, http://dx.doi.org/10.1095/biolreprod.107.066175.









