Роль глиального нейротрофического фактора в функционировании нервной системы (обзор)
Глиальный нейротрофический фактор (GDNF) — один из наиболее важных факторов выживания нейронов, способствующий дифференцировке и поддержанию различных популяций клеток центральной и периферической нервной системы. В отличие от многих других нейротрофических факторов GDNF не связывается со своим рецептором напрямую, для реализации его биологических функций необходимо присутствие корецептора, играющего роль посредника при взаимодействии с GDNF-рецептором. В качестве основного рецептора для GDNF выступает рецептор с тирозинкиназной активностью Ret, запускающий под действием GDNF последующий внутриклеточный молекулярный каскад.
Особый интерес исследователей к данному нейротрофическому фактору вызван тем, что среди других нейротрофических факторов GDNF обладает мощным нейропротективным эффектом. В связи с этим в последние годы идет активное изучение этого фактора как возможного корректора при различных нарушениях работы нервной системы, в том числе при нейродегенеративных заболеваниях.
В обзоре собрана основная информация о молекулярном строении GDNF и его рецепторов, рассмотрены механизмы реализации основных функций нейротрофического фактора, начиная с образования активного рецепторного комплекса, последующего запуска внутриклеточных сигнальных каскадов и проявления соответствующего клеточного ответа. Приведены данные публикаций, указывающие на возможность влияния GDNF на синаптогенез.
Нейротрофические факторы — полипептиды, которые регулируют развитие, поддержание, функционирование и пластичность центральной нервной системы позвоночных. Хотя первоначально эти факторы были определены как факторы выживания нейронов, они также контролируют многие другие нейронные процессы, начиная от клеточной пролиферации, дифференцировки аксонов, роста дендритов и модуляции синаптической передачи до функциональной активности нейронных ансамблей [1, 2].
Действие нейротрофических факторов заключается в модуляции биологических процессов, осуществляемых на различных уровнях. В общем виде это влияние состоит в регуляции экспрессии генов функционально значимых белков, рецепторов, медиаторов и, соответственно, во включении и/или выключении альтернативных регуляторных систем [3–5].
Один из эндогенных нейротрофических факторов, рассматривающийся как мощный терапевтический агент, — глиальный нейротрофический фактор (GDNF). Основное его действие связано с влиянием на центральную нервную систему, однако описаны его функции и в других тканях [6–8].
GDNF — необходимый фактор для нормального развития мозга в процессе эмбриогенеза, он способствует выживанию и дифференцировке различных популяций нейронов. GDNF играет важную нейропротективную роль при нейродегенеративных заболеваниях, патологиях центральной нервной системы. Многие работы указывают на его терапевтическое действие при болезни Паркинсона, ишемии головного мозга. Однако механизмы реализации действия GDNF раскрыты не до конца [9–12].
Структура глиального нейротрофического фактора
Глиальный нейротрофический фактор был впервые выделен из глиальных клеток в 1993 г. и охарактеризован как фактор выживания эмбриональных дофаминергических нейронов мозга в культуре. Позже стало ясно, что GDNF также действует как мощный нейротрофический фактор для других типов нейронов in vivo в центральной и периферической нервной системах [13, 14].
GDNF является белковой молекулой, которая содержит цистеиновый «узел» и характеризуется двумя длинными сигнальными последовательностями, образованными парами антипараллельных β-нитей [15]. Для формирования димера мономеры связываются в положении «головка–хвост». В связи с антипараллельным расположением структура GDNF имеет лево-правую симметрию, которая указывает на то, что в структуре нейротрофина присутствует симметричный участок связывания димеризованного рецептора. Структурно-функциональный анализ показал, что первые 39 аминокислот на N-конце GDNF не требуются для его биологической активности. 3D-структуры на N-конце отсутствуют. С-конец имеет решающее значение для стабильности и биологической активности GDNF, поскольку на С-конце нейротрофина расположены β-спираль и сигнальные последовательности, участвующие в связывании GDNF с GFRα1-рецептором [16–18].
Незрелая молекула GDNF состоит из 211 аминокислот, участков расщепления сигнальной последовательности и продомена. Зрелые молекулы имеют молекулярную массу 35 кДа и состоят из 134 аминокислот. В процессе созревания происходит гликозилирование белка и образование гомодимера за счет ковалентных дисульфидных связей [13]. Именно в форме гликозилированного гомодимера реализуются различные биологические функции данного нейротрофического фактора. GDNF синтезируется в виде белка-предшественника — pro-GDNF.
Установлены две формы незрелого пептида: (a)pro-GDNF и (b)pro-GDNF. Образование двух различных изоформ фермента обусловлено альтернативным сплайсингом мРНК. Показано, что (b)pro-GDNF способен индуцировать Са2+-зависимую деполяризацию нейронов [19]. Обнаружено, что изоформа (a)pro-GDNF локализуется в аппарате Гольджи, в то время как (b)pro-GDNF относится к секреторной фракции. Роль различных изоформ GDNF в настоящее время не определена. В человеческом мозге обнаружены дополнительные изоформы белка, одна из которых характерна для пациентов с болезнью Альцгеймера [20].
Ген GDNF локализуется на хромосоме 5p12-P13.1. Он содержит два экзона, один из которых кодирует зрелый белок GDNF, а также сайт расщепления, используемый при обработке белка-предшественника pro-GDNF [21, 22].
Семейство GDNF состоит из четырех членов: глиальный нейротрофический фактор, нейротурин, артемин и перзефин. Все они играют важную роль в поддержании жизнеспособности, пролиферации, дифференцировки и миграции популяций нейронов [23].
Нейротурин (neurturin, NRTN) примерно на 42% гомологичен последовательности зрелого GDNF. Доказано влияние NRTN на выживаемость дофаминергических нейронов как in vitro, так и in vivo [24–26]. Несмотря на гомологию и способность связываться с рецепторами той же группы, биологические эффекты NRTN отличаются от GDNF-опосредованных эффектов.
Перзефин (persephin, PSPN) примерно на 40% идентичен GDNF и NRTN. Как и все другие члены семейства GDNF, он поддерживает жизнедеятельность многих, в том числе дофаминергических, нейронов мозга, мотонейронов и базальных холинергических нейронов переднего мозга [27–31].
Артемин (artemin, ARTN) — самый отдаленный член семейства GDNF, он на 36% гомологичен GDNF. Показано, что артемин способствует поддержанию выживания сенсорных и симпатических нейронов в культуре. Он способен предотвратить нейропатические боли, морфологические и нейрохимические изменения в мозге животных. Однако экспрессия данного члена семейства ограничена периодом эмбрионального развития [32–37].
Рецепторы GDNF
Сигнализация GDNF опосредована связыванием с мембрано-связанным рецептором, состоящим из двух единиц. Одной из них является лиганд-связывающий компонент, специфический для семейства лигандов GDNF-корецептор GFRα, другой — рецептор с тирозинкиназной активностью Ret. Вместе они образуют функциональный блок рецепторов для связывания GDNF [38–40].
Семейство корецепторов GFRα состоит из четырех представителей (GFRα 1–4), которые определяют специфику лиганда и выступают в качестве дополнительных корецепторов для GDNF [41]. В структуре GFRα отсутствует внутриклеточный домен, поэтому данный рецептор выполняет роль передатчика сигнала к другим белкам, в частности к рецептору с тирозинкиназной активностью Ret, который в свою очередь активирует несколько внутриклеточных сигнальных каскадов [23, 42, 43].
GDNF оказывает действие не только в месте синтеза, но и дистанционно. Установлено, что нейроны способны к эндоцитозу молекул нейротрофического фактора. Поглощенный телами и проксимальными дендритами по системе ретроградного транспорта GDNF доставляется в тело и афферентные синапсы [44–47].
Каждый лиганд семейства GDNF имеет предпочтительные GFRα-корецепторы: GFRα1 — для GDNF [38, 39], GFRα2 — для нейротурина [48–51], GFRα3 — для артемина [32, 52, 53] и GFRα4 — для перзефина [54, 55].
GFRα1-рецептор в своей структуре имеет три домена (D1, D2 и D3), которые являются общими для всех млекопитающих. GFRα1 (молекулярная масса около 47 кДа) состоит из 468 аминокислот, есть три потенциальных N-связанных сайта гликозилирования. Показано, что этот белок связывается с поверхностью клетки при помощи GPI-якоря (гликозилфосфатидилинизитольный якорь). Также GFRα1 может выступать в качестве альтернативного рецептора для нейротурина с целью активации Ret. Однако связывание NRTN с GFRα1 гораздо слабее, чем с GFRα2 [38, 56–58].
В 1997 г. были определены изоформы GFRα1, образуемые в результате альтернативного сплайсинга, — GFRα1a и GFRα1b [51, 59, 60]. GFRα1a экспрессируется во всех отделах нервной системы, а GFRα1b был найден в периферических тканях [53, 59–61].
Комплекс GDNF/GFRα для дальнейшей передачи сигнала связывается с Ret, который является общим сигнальным рецептором для лигандов семейства глиального нейротрофического фактора (GFLs) (рис. 1) [42, 56, 62, 63].
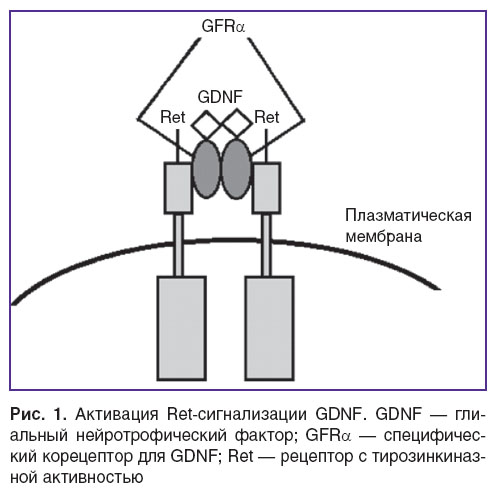 Рис. 1. Активация Ret-сигнализации GDNF. GDNF — глиальный нейротрофический фактор; GFRα — специфический корецептор для GDNF; Ret — рецептор с тирозинкиназной активностью Рис. 1. Активация Ret-сигнализации GDNF. GDNF — глиальный нейротрофический фактор; GFRα — специфический корецептор для GDNF; Ret — рецептор с тирозинкиназной активностью
|
Ген, кодирующий рецептор с тирозинкиназной активностью Ret, является протоонкогеном, который был идентифицирован в 1985 г. Установлено, что он участвует в активации перестройки ДНК [64].
Ret — трансмембранный белок, в своей внеклеточной части содержит четыре кадгерин-подобных повтора, сайт связывания кальция и цистеин-обогащенный домен. Отличие внеклеточной области молекулы Ret от других рецепторных тирозинкиназ в том, что в ней отсутствуют лейциновые повторы, иммуноглобулин и фибронектин-подобные домены, которые являются общими для многих других подобных рецепторов [65–67]. Внутриклеточная часть — типичный домен тирозинкиназы, состоящий из двух частей. Тирозинкиназный домен опосредует автофосфорилирование после активации рецептора. На основе гомологии с кадгерином кадгерин-подобные домены могут опосредовать клеточную адгезию, однако их функции в настоящее время недостаточно определены [68]. Сайт связывания кальция, расположенный между вторым и третьим кадгерин-подобными доменами, необходим для фолдинга, секреции и передачи сигнала [69–72]. Цистеин-обогащенный домен содержит 16 остатков цистеина и играет роль в связывании с корецептором GFRα [73].
Центральные функции Ret осуществляет его внутриклеточный киназный домен. Лиганд-индуцированная Ret-димеризация двух расположенных рядом каталитических доменов приводит к взаимному трансфосфорилированию и дальнейшей передаче сигнала внутриклеточным белкам [74–76].
Ret-опосредованная сигнализация GDNF включает два основных сигнальных каскада, которые способствуют выживанию клеток в различных нейрональных и ненейрональных популяциях: Ras/ERK (МАРК)- и PI3K/Akt-пути (рис. 2).
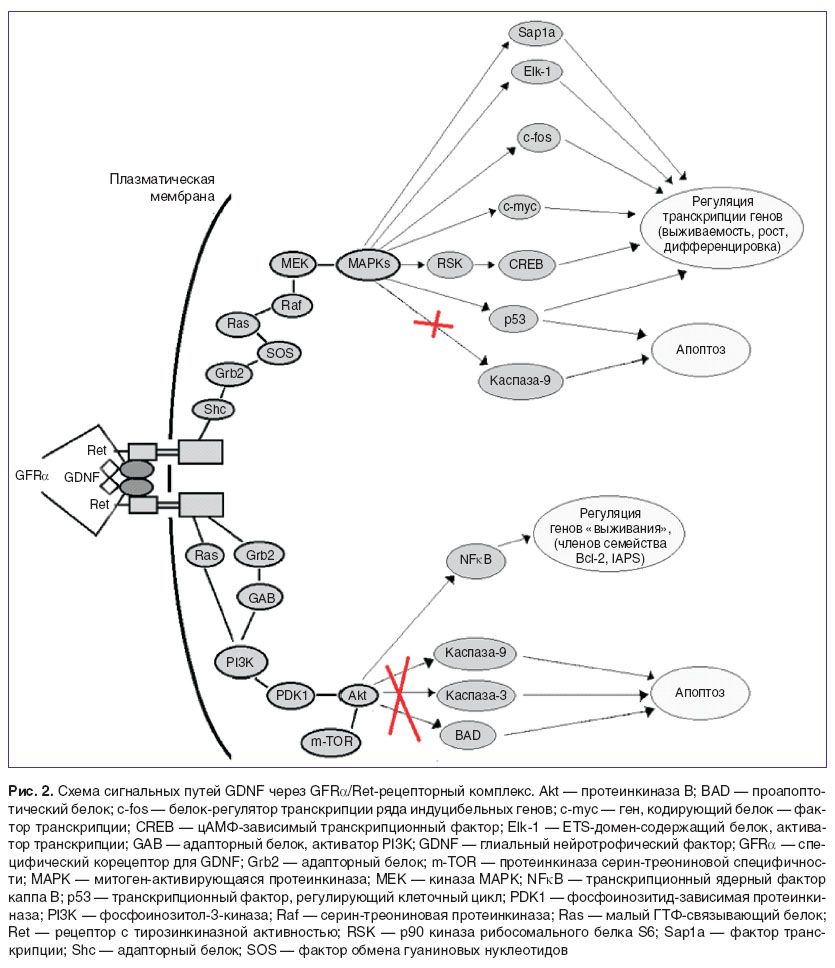 |
Рис. 2. Схема сигнальных путей GDNF через GFR/Ret-рецепторный комплекс. Akt — протеинкиназа В; BAD — проапоптотический белок; c-fos — белок-регулятор транскрипции ряда индуцибельных генов; c-myc — ген, кодирующий белок — фактор транскрипции; CREB — цАМФ-зависимый транскрипционный фактор; Elk-1 — ETS-домен-содержащий белок, активатор транскрипции; GAB — адапторный белок, активатор PI3K; GDNF — глиальный нейротрофический фактор; GFRα — специфический корецептор для GDNF; Grb2 — адапторный белок; m-TOR — протеинкиназа серин-треониновой специфичности; MAPК — митоген-активирующаяся протеинкиназа; МЕК — киназа MAPK; NFκB — транскрипционный ядерный фактор каппа В; p53 — транскрипционный фактор, регулирующий клеточный цикл; PDK1 — фосфоинозитид-зависимая протеинкиназа; PI3K — фосфоинозитол-3-киназа; Raf — серин-треониновая протеинкиназа; Ras — малый ГТФ-связывающий белок; Ret — рецептор с тирозинкиназной активностью; RSK — p90 киназа рибосомального белка S6; Sap1a — фактор транскрипции; Shc — адапторный белок; SOS — фактор обмена гуаниновых нуклеотидов |
Митоген-активированный протеинкиназный (МАРК) путь является эволюционно наиболее ранним и играет фундаментальную роль в регулировке различных клеточных процессов, включая эмбриогенез, пролиферацию, рост клеток, их дифференциацию или выживание, которые основаны на сигналах, полученных с поверхности клетки, а также о метаболическом состоянии клетки [77–79].
Запуск комплексом GDNF/GFRα/Ret MAPK-каскада приводит к активации в ядре нескольких транскрипционных факторов (в том числе с-fos и с-mус, p53, SMAD1–4, Sap1а, SP1 и Elk-1), участвующих в контроле подвижности клеток, пролиферации, дифференциации или выживания [77, 80, 81]. Известный эффект увеличения выживаемости осуществляется через фосфорилирование и активацию RSK (p90 рибосомальная S6 киназа), которая затем фосфорилирует транскрипционный фактор CREB, что приводит к активации генов «выживания» [82, 83].
Фосфоинозитол-3-киназа (PI3K) является главным регулятором выживания различных клеточных популяций [84]. Известны два основных пути активации PI3K: 1) PI3K может быть непосредственно активирована GTP-связанным Ras; 2) активация с помощью формирования Grb2/GAB1/2-комплекса. Сигнальный путь PI3K/Akt способен подавлять действие каспазы-3 и -9, апоптотический белок BAD, активировать гены «выживания» (Bcl-2 и IAPS), а также фосфорилировать транскрипционные факторы, подавляя их способность активировать апоптотические гены [85, 86].
Экспрессия GFRα в районах, лишенных Ret, и способность GDNF активировать сигнальные механизмы в клеточных линиях и первичных нейрональных культурах с низкой экспрессией Ret указывают на наличие Ret-независимого пути передачи сигнала GDNF/GFRα1. Были идентифицированы молекулы адгезии нервных клеток (NCAM) в качестве нового рецептора для GDNF в нейронах [87]. Позже было выявлено, что NCAM также является дополнительным рецептором для еще как минимум двух членов семейства лигандов GDNF — NRTN и ARTN [88].
NCAM — молекулы некадгериновой системы адгезии, трансмембранные белки, единожды пересекающие плазматическую мембрану. Внутриклеточные домены участвуют в клеточной передаче сигнала. Большая внеклеточная часть полипептидной цепи NCAM свернута в пять иммуноглобулино-подобных доменов, а также несет один или два домена, которые представляют собой повторы, встречающиеся в молекулах фибронектина [89, 90].
Связывание GFRα1 и NCAM приводит к формированию высокоаффинного рецептора для GDNF с последующей активацией цитоплазматической Src-подобной киназы Fyn и киназы фокусной адгезии FAK. Присутствие GDNF способствует адгезии клеток с помощью Fyn и FAK, при отсутствии GDNF GFRα1 ингибирует NCAM-опосредованную клеточную адгезию [87, 91]. Связывание GDNF с NCAM стимулирует миграцию шванновских клеток, рост аксонов в гиппокампе и корковых нейронах вне зависимости от наличия рецептора с тирозинкиназной активностью Ret [92, 93].
Влияние GDNF на синаптогенез
Согласно современным представлениям, минимальной функциональной единицей нервной системы является нейронная сеть. Именно на уровне нейронной сети происходят процессы консолидации памяти, обработки и передачи информации [94–96]. Каждый нейрон является частью сети и постоянно участвует в передаче информации. Суммарный сигнал, получаемый от нейронов сети, приводит к изменению мембранного потенциала и генерации потенциала действия, который в свою очередь передается другим нейронам, входящим в данный функциональный ансамбль. Особый интерес представляют сведения о роли отдельных сигнальных молекул, в частности нейротрофических факторов, в работе целостной сети.
Некоторые представители семейства GFLs участвуют не только в развитии синапсов, но и в синаптической пластичности. Показано, что GDNF способен стимулировать высвобождение нейротрансмиттера в дофаминергических нейронах среднего мозга и нервно-мышечных синапсах и, таким образом, регулировать образование и функциональные свойства синаптических окончаний. Увеличение количества кластеров пресинаптических везикул, наблюдаемое в дофаминергических нейронах среднего мозга, указывает на роль GDNF в пресинаптической дифференцировке [97, 98]. Установлено, что GDNF модулирует А-тип K+-каналов и, как следствие, возбудимость этих нейронов в культуре [99]. Также выявлено, что длительное применение GFLs индуцирует значительное увеличение числа и размера пресинаптических везикул и кластеров рецептора ацетилхолина (AchR), указывая, что члены семейства GFLs способны координировать развитие нервно-мышечных синапсов через пре- и постсинаптический механизмы [100]. Показано, что GDNF в культуре дофаминергических нейронов вызывает быстрое и обратимое повышение возбудимости нейронов. Этот эффект, по всей вероятности, опосредован подавлением К+-каналов с помощью механизма, который включает активацию МАРК. GDNF также вызывает увеличение проницаемости Ca+-каналов. Изменения в возбудимости нейронов и ионных каналов ведут к функциональной модуляции синаптической передачи. Таким образом, GDNF может рассматриваться как вещество, активно влияющее на паттерн сетевой функциональной активности нейронной сети [101, 102].
Особый интерес вызывают исследования роли GDNF и его основного рецептора в стабилизации синаптических контактов на ранних стадиях синаптогенеза. При изучении времени экспрессии и локализации GDNF и рецептора в развивающемся гиппокампе установлено, что GDNF и GFRα1 являются лиганд-индуцируемыми молекулами клеточной адгезии [103]. Иммобилизованный источник экзогенного рецептора GFRα1, имитирующего постсинаптическую локализацию, способен индуцировать дифференциацию нейронов гиппокампа. Подобные эффекты были получены для возбуждающих и тормозных нейронов гиппокампа [103]. Тем не менее актуальным остается решение вопроса, является ли этот механизм общим для всех нейрональных популяций. Показано, что развитие пресинаптической терминали, индуцированное GDNF, не зависит от Ret-опосредованных внутриклеточных механизмов и частично зависит от NCAM. Таким образом, в развитии, вероятнее всего, участвуют дополнительные эффекторные молекулярные реакции [104, 105]. Способность GDNF вызывать трансгомофильные взаимодействия между молекулами GFRα1 может рассматриваться как один из механизмов синаптогенеза. Данный механизм связан с действием как растворимых, так и мембрано-связанных молекул [106].
Исследования показали, что нейротрофические факторы GDNF и BDNF способны стимулировать промоторную активность GluR2-субъединиц AMPA-рецепторов, играющих важную роль в синаптогенезе и формировании нейронной сети, а также в синаптической пластичности, в том числе долговременной потенциации (LTP) и долговременной депрессии (LTD) [107, 108, 109], через нейронподавляющий элемент (NRSE) [110].
Заключение
GDNF играет ключевую роль в нейрогенезе, а также является необходимым фактором для поддержания жизнеспособности и функционирования нейронов. Защитное действие GDNF опосредовано его способностью блокировать апоптоз, запуская в клетке сигнальные каскады, влияющие на экспрессию генов. GDNF реализует нейротрофическую активность через формирование активного комплекса со своими рецепторами — GDNF/GFRα/Ret. Данный комплекс активизирует работу сигнальных путей MAPК и PI3K, результатом действия которых является активация транскрипционных факторов, а также подавление проапоптотических белков и каспаз.
Несмотря на это, механизмы физиологического действия GDNF, а также весь спектр его нейропротекторного потенциала до конца не выяснены. Исследование всех аспектов влияния GDNF на адаптацию нейронных сетей мозга к стресс-условиям, а также решение проблемы прохождения белка через гематоэнцефалический барьер могут дать толчок к разработке новых терапевтических стратегий и созданию лекарственных средств на основе использования данного нейротрофического фактора.
Финансирование исследования. Работа выполнена при поддержке Министерства образования и науки РФ в рамках федеральной целевой программы «Исследования и разработки по приоритетным направлениям развития научно-технического комплекса России на 2014–2020 годы». Соглашение о предоставлении субсидии №14.578.21.0094 от 24.11.2014 г. (уникальный идентификатор проекта RFMEFI57814X0094).
Конфликт интересов. У авторов нет конфликта интересов.
Литература
- Chao M.V. Neurotrophins and their receptors: a convergence point for many signalling pathways. Nat Rev Neurosci 2003; 4(4): 299–309, http://dx.doi.org/10.1038/nrn1078.
- Davies A.M. Regulation of neuronal survival and death by extracellular signals during development. EMBO J 2003; 22(11): 2537–2545, http://dx.doi.org/10.1093/emboj/cdg254.
- Гомазков О.А. Нейротрофические факторы мозга: справочно-информационное издание. CD-версия. М; 2004.
- Sakharnova T.A., Vedunova M.V., Mukhina I.V. Brain-derived neurotrophic factor (BDNF) and its role in the functioning of the central nervous system. Neurochemical Journal 2012; 6(4): 251–259, http://dx.doi.org/10.1134/s1819712412030129.
- Vedunova M.V., Mishchenko T.A., Mitroshina E.V., Mukhina I.V. TrkB-mediated neuroprotective and antihypoxic properties of brain-derived neurotrophic factor. Oxid Med Cell Longev 2015; 2015: 453901, http://dx.doi.org/10.1155/2015/453901.
- Costantini F., Shakya R. GDNF/Ret signaling and the development of the kidney. Bioessays 2006; 28(2): 117–127, http://dx.doi.org/10.1002/bies.20357.
- Naughton C.K., Jain S., Strickland A.M., Gupta A., Milbrandt J. Glial cell-line derived neurotrophic factor-mediated RET signaling regulates spermatogonial stem cell fate. Biol Reprod 2006; 74(2): 314–321, http://dx.doi.org/10.1095/biolreprod.105.047365.
- Mironov V.I., Romanov A.S., Simonov A.Y., Vedunova M.V., Kazantsev V.B. Oscillations in a neurite growth model with extracellular feedback. Neurosci Lett 2014; 570: 16–20, http://dx.doi.org/10.1016/j.neulet.2014.03.041.
- Mickiewicz A.L., Kordower J.H. GDNF family ligands: a potential future for Parkinson’s disease therapy. CNS Neurol Disord Drug Targets 2011; 10(6): 703–711, http://dx.doi.org/10.2174/187152711797247876.
- Duarte E.P., Curcio M., Canzoniero L.M., Duarte C.B. Neuroprotection by GDNF in the ischemic brain. Growth Factors 2012; 30(4): 242–257, http://dx.doi.org/10.3109/08977194.2012.691478.
- Allen S.J., Watson J.J., Shoemark D.K., Barua N.U., Patel N.K. GDNF, NGF and BDNF as therapeutic options for neurodegeneration. Pharmacol Ther 2013; 138(2): 155–175, http://dx.doi.org/10.1016/j.pharmthera.2013.01.004.
- Vedunova М.V., Sakharnova Т.А., Mitroshina E.V., Shishkina T.V., Astrakhanova T.A., Mukhina I.V. Antihypoxic and neuroprotective properties of BDNF and GDNF in vitro and in vivo under hypoxic conditions. Sovremennye tehnologii v medicine 2014; 6(4): 38–47.
- Lin L.F., Doherty D.H., Lile J.D., Bektesh S., Collins F. GDNF: a glial cell line-derived neurotrophic factor for midbrain dopaminergic neurons. Science 1993; 260(5111): 1130–1132, http://dx.doi.org/10.1126/science.8493557.
- He Z., Jiang J., Kokkinaki M., Golestaneh N., Hofmann M.C., Dym M. GDNF upregulates c-Fos transcription via the Ras/Erk1/2 pathway to promote mouse spermatogonial stem cell proliferation. Stem Cells 2008; 26(1): 266–278, http://dx.doi.org/10.1634/stemcells.2007-0436.
- Eigenbrot C., Gerber N. X-ray structure of glial cell-derived neurotrophic factor at 1.9 A resolution and implications for receptor binding. Nat Struct Biol 1997; 4(6): 435–438, http://dx.doi.org/10.1038/nsb0697-435.
- Chen Z.Y., He Z.Y., He C., Lu C.L., Wu X.F. Human glial cell-line-derived neurotrophic factor: a structure-function analysis. Biochem Biophys Res Commun 2000; 268(3): 692–696, http://dx.doi.org/10.1006/bbrc.2000.2196.
- Chen Z., He Z., He C., Lu C., Wu X. A structure-function analysis of human GDNF. Acta Biochimica et Biophysica Sinica 2000; 32(3): 243–247.
- Parkash V., Lindholm P., Peränen J., Kalkkinen N., Oksanen E., Saarma M., Leppänen V.M., Goldman A. The structure of the conserved neurotrophic factors MANF and CDNF explains why they are bifunctional. Protein Eng Des Sel 2009; 22(4): 233–241, http://dx.doi.org/10.1093/protein/gzn080.
- Lonka-Nevalaita L., Lume M., Leppänen S., Jokitalo E., Peränen J., Saarma M. Characterization of the intracellular localization, processing, and secretion of two glial cell line-derived neurotrophic factor splice isoforms. J Neurosci 2010; 30(34): 11403–11413, http://dx.doi.org/10.1523/JNEUROSCI.5888-09.2010.
- Airavaara M., Pletnikova O., Doyle M.E., Zhang Y.E., Troncoso J.C., Liu Q.R. Identification of novel GDNF isoforms and cis-antisense GDNFOS gene and their regulation in human middle temporal gyrus of Alzheimer disease. J Biol Chem 2011; 286(52): 45093–45102, http://dx.doi.org/10.1074/jbc.M111.310250.
- Schindelhauer D., Schuffenhauer S., Gasser T., Steinkasserer A., Meitinger T. The gene coding for glial cell line derived neurotrophic factor (GDNF) maps to chromosome 5p12-p13.1. Genomics 1995; 28(3): 605–607, http://dx.doi.org/10.1006/geno.1995.1202.
- Woodbury D., Schaar D.G., Ramakrishnan L., Black I.B. Novel structure of the human GDNF gene. Brain Res 1998; 803(1–2): 95–104, http://dx.doi.org/10.1016/S0006-8993(98)00627-1.
- Sariola H., Saarma M. Novel functions and signalling pathways for GDNF. J Cell Sci 2003; 116(Pt 19): 3855–3862, http://dx.doi.org/10.1242/jcs.00786.
- Kotzbauer P.T., Lampe P.A., Heuckeroth R.O., Golden J.P., Creedon D.J., Johnson E.M. Jr., Milbrandt J. Neurturin, a relative of glial-cell-line-derived neurotrophic factor. Nature 1996; 384(6608): 467–470, http://dx.doi.org/10.1038/384467a0.
- Horger B.A., Nishimura M.C., Armanini M.P., Wang L.S., Poulsen K.T., Rosenblad C., Kirik D., Moffat B., Simmons L., Johnson E. Jr., Milbrandt J., Rosenthal A., Bjorklund A., Vandlen R.A., Hynes M.A., Phillips H.S. Neurturin exerts potent actions on survival and function of midbrain dopaminergic neurons. J Neurosci 1998; 18(13): 4929–4937.
- Zihlmann K.B., Ducray A.D., Schaller B., Huber A.W., Krebs S.H., Andres R.H., Seiler R.W., Meyer M., Widmer H.R. The GDNF family members neurturin, artemin and persephin promote the morphological differentiation of cultured ventral mesencephalic dopaminergic neurons. Brain Res Bull 2005; 68(1–2): 42–53, http://dx.doi.org/10.1016/j.brainresbull.2004.10.012.
- Widenfalk J., Nosrat C., Tomac A., Westphal H., Hoffer B., Olson L. Neurturin and glial cell line-derived neurotrophic factor receptor-beta (GDNFR-beta), novel proteins related to GDNF and GDNFR-alpha with specific cellular patterns of expression suggesting roles in the developing and adult nervous system and in peripheral organs. J Neurosci 1997; 17(21): 8506–8519.
- Luukko K., Saarma M., Thesleff I. Neurturin mRNA expression suggests roles in trigeminal innervation of the first branchial arch and in tooth formation. Dev Dyn 1998; 213(2): 207–219, http://dx.doi.org/10.1002/(SICI)1097-0177(199810)213:2207::AID-AJA63.0.CO;2-K.
- Milbrandt J., de Sauvage F.J., Fahrner T.J., Baloh R.H., Leitner M.L., Tansey M.G., Lampe P.A., Heuckeroth R.O., Kotzbauer P.T., Simburger K.S., Golden J.P., Davies J.A., Vejsada R., Kato A.C., Hynes M., Sherman D., Nishimura M., Wang L.C., Vandlen R., Moffat B., Klein R.D., Poulsen K., Gray C., Garce A., Johnson E.M. Jr. Persephin, a novel neurotrophic factor related to GDNF and neurturin. Neuron 1998; 20(2): 245–253, http://dx.doi.org/10.1016/S0896-6273(00)80453-5.
- Golden J.P., DeMaro J.A., Osborne P.A., Milbrandt J., Johnson E.M. Jr. Expression of neurturin, GDNF, and GDNF family-receptor mRNA in the developing and mature mouse. Exp Neurol 1999; 158(2): 504–528, http://dx.doi.org/10.1006/exnr.1999.7127.
- Golden J.P., Milbrandt J., Johnson E.M. Jr. Neurturin and persephin promote the survival of embryonic basal forebrain cholinergic neurons in vitro. Exp Neurol 2003; 184(1): 447–455, http://dx.doi.org/10.1016/j.expneurol.2003.07.999.
- Baloh R.H., Tansey M.G., Lampe P.A., Fahrner T.J., Enomoto H., Simburger K.S., Leitner M.L., Araki T., Johnson E.M. Jr., Milbrandt J. Artemin, a novel member of the GDNF ligand family, supports peripheral and central neurons and signals through the GFRalpha3-RET receptor complex. Neuron 1998; 21(6): 1291–1302, http://dx.doi.org/10.1016/S0896-6273(00)80649-2.
- Nishino J., Mochida K., Ohfuji Y., Shimazaki T., Meno C., Ohishi S., Matsuda Y., Fujii H., Saijoh Y., Hamada H. GFR alpha3, a component of the artemin receptor, is required for migration and survival of the superior cervical ganglion. Neuron 1999; 23(4): 725–736, http://dx.doi.org/10.1016/S0896-6273(01)80031-3.
- Honma Y., Araki T., Gianino S., Bruce A., Heuckeroth R., Johnson E., Milbrandt J. Artemin is a vascular-derived neurotropic factor for developing sympathetic neurons. Neuron 2002; 35(2): 267–282, http://dx.doi.org/10.1016/S0896-6273(02)00774-2.
- Gardell L.R., Wang R., Ehrenfels C., Ossipov M.H., Rossomando A.J., Miller S., Buckley C., Cai A.K., Tse A., Foley S.F., Gong B., Walus L., Carmillo P., Worley D., Huang C., Engber T., Pepinsky B., Cate R.L., Vanderah T.W., Lai J., Sah D.W., Porreca F. Multiple actions of systemic artemin in experimental neuropathy. Nat Med 2003; 9(11): 1383–1389, http://dx.doi.org/10.1038/nm944.
- Bennett D.L., Boucher T.J., Michael G.J., Popat R.J., Malcangio M., Averill S.A., Poulsen K.T., Priestley J.V., Shelton D.L., McMahon S.B. Artemin has potent neurotrophic actions on injured C-fibres. J Peripher Nerv Syst 2006; 11(4): 330–345, http://dx.doi.org/10.1111/j.1529-8027.2006.00106.x.
- Thornton P., Hatcher J.P., Robinson I., Sargent B., Franzén B., Martino G., Kitching L., Glover C.P., Anderson D., Forsmo-Bruce H., Low C.P., Cusdin F., Dosanjh B., Williams W., Steffen A.C., Thompson S., Eklund M., Lloyd C., Chessell I., Hughes J. Artemin-GFRα3 interactions partially contribute to acute inflammatory hypersensitivity. Neurosci Lett 2013; 545: 23–28, http://dx.doi.org/10.1016/j.neulet.2013.04.007.
- Jing S., Wen D., Yu Y., Holst P.L., Luo Y., Fang M., Tamir R., Antonio L., Hu Z., Cupples R., Louis J.C., Hu S., Altrock B.W., Fox G.M. GDNF-induced activation of the ret protein tyrosine kinase is mediated by GDNFR-alpha, a novel receptor for GDNF. Cell 1996; 85(7): 1113–1124, http://dx.doi.org/10.1016/S0092-8674(00)81311-2.
- Treanor J.J., Goodman L., de Sauvage F., Stone D.M., Poulsen K.T., Beck C.D., Gray C., Armanini M.P., Pollock R.A., Hefti F., Phillips H.S., Goddard A., Moore M.W., Buj-Bello A., Davies A.M., Asai N., Takahashi M., Vandlen R., Henderson C.E., Rosenthal A. Characterization of a multicomponent receptor for GDNF. Nature 1996; 382(6586): 80–83, http://dx.doi.org/10.1038/382080a0.
- Aron L. Genetic analysis of dopaminergic neuron survival GDNF/Ret signaling and the Parkinson’s disease-associated gene DJ-1 [dissertation]. München: Ludwig-Maximilians-Universität München; 2009.
- Serra M.P., Quartu M., Mascia F., Manca A., Boi M., Pisu M.G., Lai M.L., Del Fiacco M. Ret, GFRalpha-1, GFRalpha-2 and GFRalpha-3 receptors in the human hippocampus and fascia dentate. Int J Dev Neurosci 2005; 23: 425–438, http://dx.doi.org/10.1016/j.ijdevneu.2005.05.003.
- Airaksinen M.S., Saarma M. The GDNF family: signalling, biological functions and therapeutic value. Nat Rev Neurosci 2002; 3(5): 383–394, http://dx.doi.org/10.1038/nrn812.
- Lucini C., Facello B., Maruccio L., Langellotto F., Sordino P., Castaldo L. Distribution of glial cell line-derived neurotrophic factor receptor alpha-1 in the brain of adult zebrafish. J Anat 2010; 217(2): 174–185, http://dx.doi.org/10.1111/j.1469-7580.2010.01254.x.
- Tomac A., Widenfalk J., Lin L.F., Kohno T., Ebendal T., Hoffer B.J., Olson L. Retrograde axonal transport of glial cell line-derived neurotrophic factor in the adult nigrostriatal system suggests a trophic role in the adult. Proc Natl Acad Sci USA 1995; 92(18): 8274–8278, http://dx.doi.org/10.1073/pnas.92.18.8274.
- Leitner M.L., Molliver D.C., Osborne P.A., Vejsada R., Golden J.P., Lampe P.A., Kato A.C., Milbrandt J., Johnson E.M. Jr. Analysis of the retrograde transport of glial cell line-derived neurotrophic factor (GDNF), neurturin, and persephin suggests that in vivo signaling for the GDNF family is GFRalpha coreceptor-specific. J Neurosci 1999; 19(21): 9322–9331.
- Rind H.B., Butowt R., von Bartheld C.S. Synaptic targeting of retrogradely transported trophic factors in motoneurons: comparison of glial cell line-derived neurotrophic factor, brain-derived neurotrophic factor, and cardiotrophin-1 with tetanus toxin. J Neurosci 2005; 25(3): 539–549, http://dx.doi.org/10.1523/jneurosci.4322-04.2005.
- Tsui C.C., Pierchala B.A. The differential axonal degradation of Ret accounts for cell-type-specific function of glial cell line-derived neurotrophic factor as a retrograde survival factor. J Neurosci 2010; 30: 5149–5158, http://dx.doi.org/10.1523/JNEUROSCI.5246-09.2010.
- Baloh R.H., Tansey M.G., Golden J.P., Creedon D.J., Heuckeroth R.O., Keck C.L., Zimonjic D.B., Popescu N.C., Johnson E.M. Jr., Milbrandt J. TrnR2, a novel receptor that mediates neurturin and GDNF signaling through Ret. Neuron 1997; 18(5): 793–802, http://dx.doi.org/10.1016/S0896-6273(00)80318-9.
- Buj-Bello A., Adu J., Piñón L.G., Horton A., Thompson J., Rosenthal A., Chinchetru M., Buchman V.L., Davies A.M. Neurturin responsiveness requires a GPI-linked receptor and the Ret receptor tyrosine kinase. Nature 1997; 387(6634): 721–724, http://dx.doi.org/10.1038/42729.
- Klein R.D., Sherman D., Ho W.H., Stone D., Bennett G.L., Moffat B., Vandlen R., Simmons L., Gu Q., Hongo J.A., Devaux B., Poulsen K., Armanini M., Nozaki C., Asai N., Goddard A., Phillips H., Henderson C.E., Takahashi M., Rosenthal A. A GPI-linked protein that interacts with Ret to form a candidate neurturin receptor. Nature 1997; 387: 717–721, http://dx.doi.org/10.1038/42722.
- Sanicola M., Hession C., Worley D., Carmillo P., Ehrenfels C., Walus L., Robinson S., Jaworski G., Wei H., Tizard R., Whitty A., Pepinsky R.B., Cate R.L. Glial cell line-derived neurotrophic factor-dependent RET activation can be mediated by two different cell-surface accessory proteins. Proc Natl Acad Sci USA 1997; 94(12): 6238–6243, http://dx.doi.org/10.1073/pnas.94.12.6238.
- Naveilhan P., Baudet C., Mikaels A., Shen L., Westphal H., Ernfors P. Expression and regulation of GFRalpha3, a glial cell line-derived neurotrophic factor family receptor. Proc Natl Acad Sci USA 1998; 95(3): 1295–1300, http://dx.doi.org/10.1073/pnas.95.3.1295.
- Widenfalk J., Tomac A., Lindqvist E., Hoffer B., Olson L. GFRalpha-3, a protein related to GFRalpha-1, is expressed in developing peripheral neurons and ensheathing cells. Eur J Neurosci 1998; 10(4): 1508–1517, http://dx.doi.org/10.1046/j.1460-9568.1998.00192.x.
- Enokido Y., de Sauvage F., Hongo J.A., Ninkina N., Rosenthal A., Buchman V.L., Davies A.M. GFRalpha-4 and the tyrosine kinase Ret form a functional receptor complex for persephin. Curr Biol 1998; 8(18): 1019–1022, http://dx.doi.org/10.1016/s0960-9822(07)00422-8.
- Lindahl M., Poteryaev D., Yu L., Arumae U., Timmusk T., Bongarzone I., Aiello A., Pierotti M.A., Airaksinen M.S., Saarma M. Human glial cell line-derived neurotrophic factor receptor alpha 4 is the receptor for persephin and is predominantly expressed in normal and malignant thyroid medullary cells. J Biol Chem 2001; 276(12): 9344–9351, http://dx.doi.org/10.1074/jbc.m008279200.
- Creedon D.J., Tansey M.G., Baloh R.H., Osborne P.A., Lampe P.A., Fahrner T.J., Heuckeroth R.O., Milbrandt J., Johnson E.M. Jr. Neurturin shares receptors and signal transduction pathways with glial cell line-derived neurotrophic factor in sympathetic neurons. Proc Natl Acad Sci USA 1997; 94(13): 7018–7023, http://dx.doi.org/10.1073/pnas.94.13.7018.
- Suvanto P., Wartiovaara K., Lindahl M., Arumae U., Moshnyakov M., Horelli-Kuitunen N., Airaksinen M.S., Palotie A., Sariola H., Saarma M. Cloning, mRNA distribution and chromosomal localisation of the gene for glial cell line-derived neurotrophic factor receptor beta, a homologue to GDNFR-alpha. Hum Mol Genet 1997; 6(8): 1267–1273, http://dx.doi.org/10.1093/hmg/6.8.1267.
- Airaksinen M.S., Titievsky A., Saarma M. GDNF family neurotrophic factor signaling: four masters, one servant? Mol Cell Neurosci 1999; 13(5): 313–325, http://dx.doi.org/10.1006/mcne.1999.0754.
- Dey B.K., Wong Y.W., Too H.P. Cloning of a novel murine isoform of the glial cell line-derived neurotrophic factor receptor. Neuroreport 1998; 9(1): 37–42, http://dx.doi.org/10.1097/00001756-199801050-00008.
- Shefelbine S.E., Khorana S., Schultz P.N., Huang E., Thobe N., Hu Z.J., Fox G.M., Jing S., Cote G.J., Gagel R.F. Mutational analysis of the GDNF/RET-GDNFR alpha signaling complex in a kindred with vesicoureteral reflux. Hum Genet 1998; 102(4): 474–478, http://dx.doi.org/10.1007/s004390050724.
- Yoong L.F., Peng Z.N., Wan G., Too H.P. Tissue expression of alternatively spliced GFRalpha1, NCAM and RET isoforms and the distinct functional consequence of ligand-induced activation of GFRalpha1 isoforms. Brain Res Mol Brain Res 2005; 139(1): 1–12, http://dx.doi.org/10.1016/j.molbrainres.2005.05.016.
- Baloh R.H., Enomoto H., Johnson E.M. Jr., Milbrandt J. The GDNF family ligands and receptors — implications for neural development. Curr Opin Neurobiol 2000; 10(1): 103–110, http://dx.doi.org/10.1016/s0959-4388(99)00048-3.
- Wang X. Structural studies of GDNF family ligands with their receptors — Insights into ligand recognition and activation of receptor tyrosine kinase RET. Biochim Biophys Acta 2013; 1834(10): 2205–2212, http://dx.doi.org/10.1016/j.bbapap.2012.10.008.
- Takahashi M., Ritz J., Cooper G.M. Activation of a novel human transforming gene, ret, by DNA rearrangement. Cell 1985; 42(2): 581–588, http://dx.doi.org/10.1016/0092-8674(85)90115-1.
- Iwamoto T., Taniguchi M., Asai N., Ohkusu K., Nakashima I., Takahashi M. cDNA cloning of mouse ret proto-oncogene and its sequence similarity to the cadherin superfamily. Oncogene 1993; 8(4): 1087–1091.
- Schneider R. The human protooncogene ret: a communicative cadherin? Trends Biochem Sci 1992; 17(11): 468–469, http://dx.doi.org/10.1016/0968-0004(92)90490-z.
- Traugott A.L., Moley J.F. The RET protooncogene. Cancer Treat Res 2010; 153: 303–319, http://dx.doi.org/10.1007/978-1-4419-0857-5_17.
- Anders J., Kjar S., Ibáсez C.F. Molecular modeling of the extracellular domain of the RET receptor tyrosine kinase reveals multiple cadherin-like domains and a calcium-binding site. J Biol Chem 2001; 276(38): 35808–35817, http://dx.doi.org/10.1074/jbc.m104968200.
- Nozaki C., Asai N., Murakami H., Iwashita T., Iwata Y., Horibe K., Klein R.D., Rosenthal A., Takahashi M. Calcium-dependent Ret activation by GDNF and neurturin. Oncogene 1998; 16(3): 293–299, http://dx.doi.org/10.1038/sj.onc.1201548.
- van Weering D.H., Bos J.L. Signal transduction by the receptor tyrosine kinase Ret. Recent Results Cancer Res 1998; 154: 271–281, http://dx.doi.org/10.1007/978-3-642-46870-4_18.
- van Weering D.H., Moen T.C., Braakman I., Baas P.D., Bos J.L. Expression of the receptor tyrosine kinase Ret on the plasma membrane is dependent on calcium. J Biol Chem 1998; 273(20): 12077–12081, http://dx.doi.org/10.1074/jbc.273.20.12077.
- Ibáñez С .F. Structure and physiology of the RET receptor tyrosine kinase. Cold Spring Harb Perspect Biol 2013 года; 5(2): a009134, http://dx.doi.org/10.1101/cshperspect.a009134.
- Runeberg-Roos P., Saarma M. Neurotrophic factor receptor RET: structure, cell biology, and inherited diseases. Ann Med 2007; 39(8): 572–580, http://dx.doi.org/10.1080/07853890701646256.
- Liu X., Vega Q.C., Decker R.A., Pandey A., Worby C.A., Dixon J.E. Oncogenic RET receptors display different autophosphorylation sites and substrate binding specificities. J Biol Chem 1996; 271(10): 5309–5312, http://dx.doi.org/10.1074/jbc.271.10.5309.
- Kawamoto Y., Takeda K., Okuno Y., Yamakawa Y., Ito Y., Taguchi R., Kato M., Suzuki H., Takahashi M., Nakashima I. Identification of RET autophosphorylation sites by mass spectrometry. J Biol Chem 2004; 279(14): 14213–14224, http://dx.doi.org/10.1074/jbc.m312600200.
- Lemmon M.A., Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell 2010; 141(7): 1117–1134, http://dx.doi.org/10.1016/j.cell.2010.06.011.
- Dhillon A.S., Hagan S., Rath O., Kolch W. MAP kinase signalling pathways in cancer. Oncogene 2007; 26(22): 3279–3290, http://dx.doi.org/10.1038/sj.onc.1210421.
- Krishna M., Narang H. The complexity of mitogen-activated protein kinases (MAPKs) made simple. Cell Mol Life Sci 2008; 65(22): 3525–3544, http://dx.doi.org/10.1007/s00018-008-8170-7.
- Yang S.H., Sharrocks A.D., Whitmarsh A.J. MAP kinase signalling cascades and transcriptional regulation. Gene 2013; 513(1): 1–13, http://dx.doi.org/10.1016/j.gene.2012.10.033.
- Raman M., Chen W., Cobb M.H. Differential regulation and properties of MAPKs. Oncogene 2007; 26(22): 3100–3112, http://dx.doi.org/10.1038/sj.onc.1210392.
- Turjanski A.G., Vaqué J.P., Gutkind J.S. MAP kinases and the control of nuclear events. Oncogene 2007; 26(22): 3240–3253, http://dx.doi.org/10.1038/sj.onc.1210415.
- Reichardt L.F. Neurotrophin-regulated signalling pathways. Philos Trans R Soc Lond B Biol Sci 2006; 361(1473): 1545–1564, http://dx.doi.org/10.1098/rstb.2006.1894.
- Ron D., Janak P.H. GDNF and addiction. Rev Neurosci 2005; 16(4): 277–285, http://dx.doi.org/10.1515/revneuro.2005.16.4.277.
- Manning B.D., Cantley L.C. AKT/PKB signaling: navigating downstream. Cell 2007; 129(7): 1261–1274, http://dx.doi.org/10.1016/j.cell.2007.06.009.
- Kim D., Chung J. Akt: versatile mediator of cell survival and beyond. J Biochem Mol Biol 2002; 35(1): 106–115, http://dx.doi.org/10.5483/bmbrep.2002.35.1.106.
- Datta S.R., Brunet A., Greenberg M.E. Cellular survival: a play in three Akts. Genes Dev 1999; 13(22): 2905–2927, http://dx.doi.org/10.1101/gad.13.22.2905.
- Paratcha G., Ledda F., Ibáсez C.F. The neural cell adhesion molecule NCAM is an alternative signaling receptor for GDNF family ligands. Cell 2003; 113(7): 867–879, http://dx.doi.org/10.1016/S0092-8674(03)00435-5.
- Schmutzler B.S., Roy S., Pittman S.K., Meadows R.M., Hingtgen C.M. Ret-dependent and Ret-independent mechanisms of Gfl-induced sensitization. Mol Pain 2011; 7: 22, http://dx.doi.org/10.1186/1744-8069-7-22.
- Gegelashvili G., Bock E., Schousboe A., Linnemann D. Two types of amyloid precursor protein (APP) mRNA in rat glioma cell lines: upregulation via a cyclic AMP-dependent pathway. Brain Res Mol Brain Res 1996; 37(1–2): 151–156, http://dx.doi.org/10.1016/0169-328x(95)00302-9.
- Owczarek S., Kristiansen L.V., Hortsch M., Walmod P.S. Cell adhesion molecules of the NCAM family and their roles at synapses. The Sticky Synapse 2009; 265–299, http://dx.doi.org/10.1007/978-0-387-92708-4_13.
- Ibáñez C.F. Beyond the cell surface: new mechanisms of receptor function. Biochem Biophys Res Commun 2010; 396(1): 24–27, http://dx.doi.org/10.1016/j.bbrc.2010.01.136.
- Canty A.J., Dietze J., Harvey M., Enomoto H., Milbrandt J., Ibáñez C.F. Regionalized loss of parvalbumin interneurons in the cerebral cortex of mice with deficits in GFRalpha1 signaling. J Neurosci 2009; 29(34): 10695–10705, http://dx.doi.org/10.1523/JNEUROSCI.2658-09.2009.
- Sjöstrand D., Ibáсez C.F. Insights into GFRalpha1 regulation of neural cell adhesion molecule (NCAM) function from structure-function analysis of the NCAM/GFRalpha1 receptor complex. J Biol Chem 2008; 283(20): 13792–13898, http://dx.doi.org/10.1074/jbc.M800283200.
- Yuste R. From the neuron doctrine to neural networks. Nat Rev Neurosci 2015; 16(8): 487–497, http://dx.doi.org/10.1038/nrn3962.
- Schlingloff D., Káli S., Freund T.F., Hájos N., Gulyás A.I. Mechanisms of sharp wave initiation and ripple generation. J Neurosci 2014; 20(34): 11385–11398, http://dx.doi.org/10.1523/JNEUROSCI.0867-14.2014.
- Tong M.T., Peace S.T., Cleland T.A. Properties and mechanisms of olfactory learning and memory. Front Behav Neurosci 2014; 8: 238, http://dx.doi.org/10.3389/fnbeh.2014.00238.
- Bourque M.J., Trudeau L.E. GDNF enhances the synaptic efficacy of dopaminergic neurons in culture. Eur J Neurosci 2000; 12(9): 3172–3180, http://dx.doi.org/10.1046/j.1460-9568.2000.00219.x.
- Nguyen Q.T., Parsadanian A.Sh., Snider W.D., Lichtman J.W. Hyperinnervation of neuromuscular junctions caused by GDNF overexpressionin muscle. Science 1998; 279: 1725–1729, http://dx.doi.org/10.1126/science.279.5357.1725
- Yang F., Feng L., Zheng F., Johnson S.W., Du J., Shen L., Wu C.P., Lu B. GDNF acutely modulates excitability and A-type K(+) channels in midbrain dopaminergic neurons. Nat Neurosci 2001; 4(11): 1071–1078, http://dx.doi.org/10.1038/nn734.
- Wang C.Y., Yang F., He X.P., Je H.S., Zhou J.Z., Eckermann K., Kawamura D., Feng L., Shen L., Lu B. Regulation of neuromuscular synapse development by glial cell line-derived neurotrophic factor and neurturin. J Biol Chem 2002; 277(12): 10614–10625, http://dx.doi.org/10.1074/jbc.M106116200.
- Wang J., Chen G., Lu B., Wu C.P. GDNF acutely potentiates Ca2+ channels and excitatory synaptic transmission in midbrain dopaminergic neurons. Neurosignals 2003; 12(2): 78–88, http://dx.doi.org/10.1159/000071817.
- Weiss J.L., Burgoyne R.D. Sense and sensibility in the regulation of voltage-gated Ca2+ channels. Trends Neurosci 2002; 25(10): 489–491, http://dx.doi.org/10.1016/S0166-2236(02)02247-6.
- Ledda F., Paratcha G., Sandoval-Guzmán T., Ibáñez C.F. GDNF and GFRalpha1 promote formation of neuronal synapses by ligand-induced cell adhesion. Nat Neurosci 2007; 10(3): 293–300, http://dx.doi.org/10.1038/nn1855.
- Rutishauser U. Polysialic acid in the plasticity of the developing and adult vertebrate nervous system. Nat Rev Neurosci 2008; 9(1): 26–35, http://dx.doi.org/10.1038/nrn2285.
- Võikar V., Rossi J., Rauvala H., Airaksinen M.S. Impaired behavioural flexibility and memory in mice lacking GDNF family receptor alpha2. Eur J Neurosci 2004; 20(1): 308–312, http://dx.doi.org/10.1111/j.1460-9568.2004.03475.x.
- Paratcha G., Ledda F. GDNF and GFRalpha: a versatile molecular complex for developing neurons. Trends Neurosci 2008; 31(8): 384–391, http://dx.doi.org/10.1016/j.tins.2008.05.003.
- Choi D.W. Excitotoxic cell death. J Neurobiol 1992; 23(9): 1261–1276, http://dx.doi.org/10.1002/neu.480230915.
- Lipton S.A., Rosenberg P.A. Excitatory amino acids as a final common pathway for neurologic disorders. N Engl J Med 1994; 330(9): 613–622, http://dx.doi.org/10.1056/nejm199403033300907.
- Vedunova M., Sakharnova T., Mitroshina E., Perminova M., Pimashkin A., Zakharov Yu., Dityatev A., Mukhina I. Seizure-like activity in hyaluronidase-treated dissociated hippocampal cultures. Front Cell Neurosci 2013; 7: 149, http://dx.doi.org/10.3389/fncel.2013.00149.
- Brené S., Messer C., Okado H., Hartley M., Heinemann S.F., Nestler E.J. Regulation of GluR2 promoter activity by neurotrophic factors via a neuron-restrictive silencer element. Eur J Neurosci 2000; 12(5): 1525–1533, http://dx.doi.org/10.1046/j.1460-9568.2000.00040.x.









