Новые представления о механизмах действия доксорубицина и озона на злокачественные клетки печени
Цель исследования — экспериментально подтвердить схожесть пусковых механизмов действия озона и доксорубицина на культуре злокачественных клеток печени человека.
Материалы и методы. В экспериментах на культуральной среде изучали влияние химиопрепарата доксорубицина, озона и их сочетаний на злокачественные клетки печени (SK-HEP-1).
Результаты. Установлено сходное повышение показателей свободнорадикального окисления при действии озона и доксорубицина как изолированно, так и в сочетании. Их введение во всех вариантах повышало также содержание фермента каспазы-3, при этом уровни каспазы-3 были существенно выше при введении доксорубицина. Полученные результаты показывают новые механизмы влияния озона и доксорубицина на жизнеспособность и морфологические изменения в злокачественных клетках печени: активация свободнорадикального окисления вызывает в этих клетках изменения как некротические, так и апоптотические — через увеличение количества каспаз.
В проведенных ранее экспериментах [1] на лабораторных животных (крысах), которым перевивался штамм рака молочной железы, исследовалось влияние на опухоль химиопрепарата доксорубицина, озона и их комбинированного введения. Полученные результаты продемонстрировали, что сочетанное воздействие низких терапевтических концентраций озона в составе озонированного физиологического раствора и доксорубицина оказывало наиболее выраженный деструктивный эффект на опухоль. Использование озонированного физиологического раствора потенцировало противоопухолевую активность доксорубицина, что проявлялось в выраженном угнетении митотической активности опухолевых клеток и снижении числа их жизнеспособных элементов. Исследование ИК-спектров тканей опухоли, печени, легких, мозга животных-опухоленосителей [2] также подтвердило более высокий терапевтический эффект сочетанного действия доксорубицина и озона.
Продолжением исследований явились эксперименты на культуре нормальных клеток печени Chang liver и злокачественных клеток печени SK-HEP-1 человека [3]. Установлено, что введение озона в культуральную среду оказывает сходный с доксорубицином выраженный цитостатический эффект на жизнеспособность клеток, что подтверждалось морфологическими данными о необратимых изменениях в структуре клеточных элементов некротического или апоптотического происхождения. Несмотря на доказательный материал по жизнеспособности и патоморфозу злокачественных клеток под действием анализируемых факторов, внутриклеточный механизм запуска гибели клеток остается неясным. Про доксорубицин известно, что это цитостатик антрациклинового ряда с антимитотическим и антипролиферативным действием. Механизм действия препарата объясняют его реакцией с ДНК, образованием свободных радикалов и прямым воздействием на мембраны клеток с подавлением синтеза нуклеиновых кислот. В плане активации свободнорадикальных реакций действие доксорубицина может быть сопоставимо с действием озона как сильного окислителя, регулирующего про- и антиоксидантный баланс [4–6].
Другим возможным механизмом, запускающим гибель клеток, является ферментативный путь с участием каспаз. Каспазы — это семейство аспартатспецифических цистеиновых протеаз, они присутствуют во всех клетках, взаимодействие этих протеаз с олигомерными рецепторами ведет к их активации. Активные каспазы могут запускать протеолитический каскад, расщепляющий белки, необходимые для выживания. Конечным итогом сигнального пути является активация контролируемой гибели клеток — апоптоз. Каспазы, вовлеченные в апоптоз, делятся на инициаторные и эффекторные. Одной из эффекторных каспаз является каспаза-3, расщепляющая субстрат на карбоксильном конце по остаткам аспартата. Ингибирование процесса апоптоза может приводить к развитию онкологических заболеваний [7].
Исходя из результатов собственных исследований и данных литературы можно предположить сопоставимость пусковых механизмов действия озона и доксорубицина внутри клеток.
Цель исследования — экспериментально подтвердить схожесть пусковых механизмов действия озона и доксорубицина на культуре злокачественных клеток печени человека.
Материалы и методы. Эксперименты проводили на культивированных клетках аденокарциномы печени человека SK-HEP-1, морфология — эпителиоподобная, культивирование осуществляли в среде «Игла МЕМ» с солями «Эрла» («ПанЭко», Россия) с добавлением 10% сыворотки эмбриональной телячьей («ПанЭко», Россия) и 1% заменимых аминокислот («ПанЭко», Россия), оптимальная плотность — (2,0–4,0)·106 кл./см2. Поддержание жизнеспособности клеток осуществлялось в СО2-инкубаторе при 5% содержании СО2. После 3–5 пассажей клетки рассаживали на 48- или 6-луночные планшеты. При достижении 60% монослоя среду, в которой клетки выращивались, заменяли на испытуемые среды. Первая среда готовилась добавлением химиопрепарата доксорубицина в дозе 0,004 мг; вторая — введением 150 мл кислорода; третья — введением 150 мл озоно-кислородной смеси с концентрацией озона 25 мг/л; четвертая — введением 150 мл кислорода и 0,004 мг доксорубицина; пятая — введением 150 мл озоно-кислородной смеси и 0,004 мг доксорубицина. Дозы доксорубицина и озона определены в эксперименте на животных при оценке патоморфоза злокачественной опухоли [1].
При озонировании культуральной среды озоно-кислородная газовая смесь поступала со скоростью 1 л/мин в течение 5 мин из генератора озона («Квазар», Россия). Через 48 ч культивирования клеточная среда убиралась, клетки промывались полифосфатным буфером PBS (pH=7,4) и заливались 250 мл смеси Версен (0,02%):трипсин (0,25%) (3:1). Через 10 мин инкубации в СО2-инкубаторе клетки пипетировали и добавляли в каждую лунку по 250 мл 8% формальдегида. После этого подсчитывали количество клеток на автоматическом анализаторе Septer (Мillipore, Великобритания).
Для проведения анализа на активность свободнорадикального окисления клеточную суспензию трижды промывали забуференным физиологическим раствором и замораживали при –20°. Перед началом исследования проводили ее размораживание и гомогенизацию. Интенсивность свободнорадикального окисления оценивали по параметрам индуцированной железом и перекисью водорода хемилюминесценции на аппарате БХЛ-07 (Н. Новгород, Россия): Imax — максимальная интенсивность свечения, S — cветосумма хемилюминесценции за 30 с [8]. Содержание продуктов перекисного окисления липидов — первичных диеновых конъюгатов (ДК), триеновых конъюгатов (ТК), конечных оснований Шиффа (ОШ) определяли в гептан-изопропанольных фракциях по методу И.А. Волчегорского [9]. Количество фермента каспазы-3 оценивали методом иммуноферментного анализа Human Caspase-3 Instant ELISA (ThermoFisher Scientific, США) и рассчитывали на количество клеток в мл (содержание клеток — 5·106/мл).
Полученные результаты были обработаны с помощью пакета прикладных программ Biostat и представлены в виде М±σ, где М — среднее арифметическое, σ — среднеквадратичное отклонение. Достоверность различий средних определяли по t-критерию Стьюдента. Различия считали статистически значимыми при уровне значимости p<0,05.
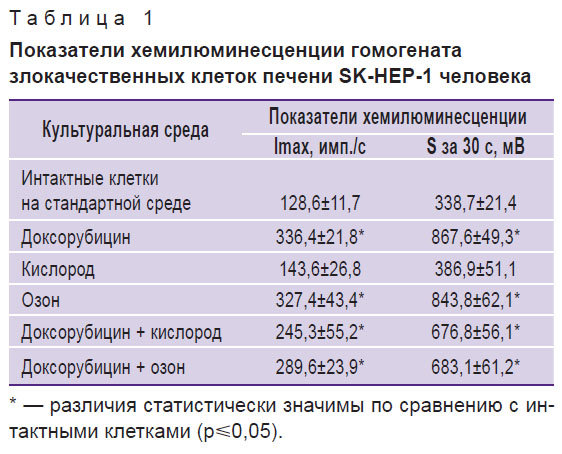
Результаты. Введение в культуральную среду для выращивания клеток аденокарциномы печени человека SK-HEP-1 доксорубицина (табл. 1) сопровождалось статистически значимым повышением уровней показателей Imax — в 2,7 раза и S — в 2,5 раза, что указывало на активацию свободнорадикальных реакций под действием цитостатика. Озонирование культуральной среды сходным с доксорубицином образом активировало свободнорадикальные реакции и проявлялось увеличением Imax в 2,6 раза и S — в 2,5 раза. Сочетанное введение в культуральную среду доксорубицина и кислорода, а также доксорубицина и озона статистически значимо не снижало высокие уровни показателей Imax и S, характерные для среды с доксорубицином, и составляло для Imax 1,8 и 2,2 раза, а для S — 2,1 и 2,5 раза соответственно.
|
Таблица 1. Показатели хемилюминесценции гомогената злокачественных клеток печени SK-HEP-1 человека |
Таким образом, предположение о том, что одним из механизмов, вызывающих снижение жизнеспособности злокачественных клеток печени и их морфологические изменения [3], являются свободнорадикальные реакции, запускаемые как окислителем, так и токсичным соединением доксорубицином, подтверждалось. Дополнительным доказательством этому явились данные по изучению уровней продуктов перекисного окисления липидов (табл. 2).
|
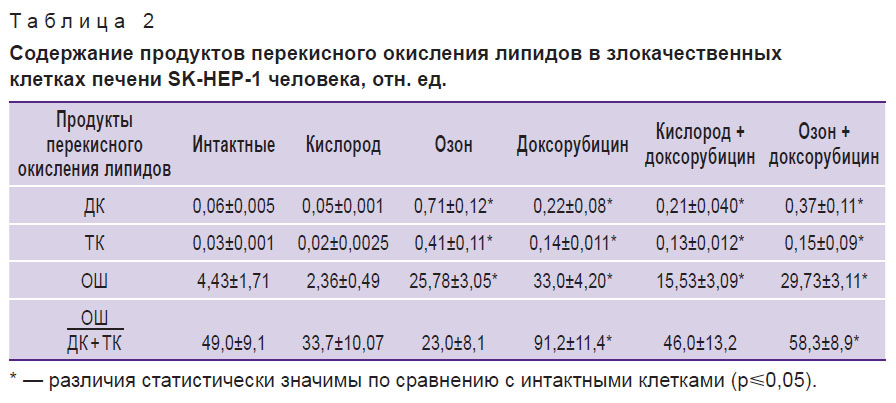
Таблица 2. Содержание продуктов перекисного окисления липидов в злокачественных клетках печени SK-HEP-1 человека |
Самые выраженные изменения в показателях, характеризующих активность перекисного окисления липидов в культуре злокачественных клеток, наблюдались в клетках, находящихся в среде с доксорубицином. Уровни ДК в этих клетках увеличивались почти в 4 раза, ТК — в 4,5 раза, ОШ — в 4 раза. Коэффициент ОШ/(ДК+ТК), представляющий собой количественное отношение конечных продуктов к первичным, повысился в 2 раза, что свидетельствовало о сдвиге реакций в сторону накопления жестких токсичных ОШ, вызывающих повреждение клеточных мембран. Озонирование культуральной среды статистически значимо увеличивало в клетках содержание ДК в 11,8 раза, ТК — в 13,3 раза, ОШ — в 4,5 раза, коэффициент ОШ/(ДК+ТК) снижался по сравнению с исходным уровнем в 2 раза, что свидетельствовало о преобладании первичных продуктов перекисного окисления липидов и, следовательно, об активно продолжающемся процессе на этапах инициации. Сочетанное введение доксорубицина с кислородом и доксорубицина с озоном практически не вызывало различий по уровням ДК и ТК, в то время как для уровней ОШ и коэффициента ОШ/(ДК+ТК) (см. табл. 2) более значимые различия наблюдались при сочетании доксорубицина и озона. Высокие уровни ОШ и коэффициента ОШ/ДК+ТК свидетельствуют о накоплении конечных продуктов перекисного окисления, которые могут повреждать клетки, что может отражаться на показателях жизнеспособности и морфологии клеток.
Помимо повреждающего действия свободнорадикального окисления на внутриклеточные структуры нельзя исключать губительного действия ферментов, вызывающих апоптоз. Одним из таких ферментов является каспаза-3. В нашем исследовании оценивалось содержание данного фермента в гомогенате клеток, содержащихся на культуральных средах, в которые вводили доксорубицин, озон, кислород и их сочетания. Самая высокая концентрация каспазы-3 отмечалась при воздействии на злокачественные клетки доксорубицином (4,76±0,06 пг/мл), она в 30 раз превышала содержание фермента в клетках интактной серии (<0,16 пг/мл). Данный факт явился подтверждением высокой апоптотической активности доксорубицина, выявляемой при морфологических исследованиях. Озонирование среды для культивирования клеток увеличивало количество каспазы в 11 раз (1,82±0,01 пг/мл). При смене среды для выращивания клеток на среду, содержащую доксорубицин и озон, количество каспазы увеличивалось в 13 раз (2,04±0,03 пг/мл), а на среду, содержащую доксорубицин и кислород, — в 16,6 раза (2,67±0,02 пг/мл).
Полученные результаты свидетельствуют о том, что основной причиной запуска апоптотического процесса в наших экспериментах со злокачественными клетками, которые содержатся в средах, подвергшихся воздействию доксорубицина или окислителей, служит повышение уровней каспазы-3. В свою очередь увеличение концентрации каспазы, скорее всего, является следствием активации свободнорадикального окисления под действием как доксорубицина, так и озона.
Заключение. Введение в культуральную среду для выращивания клеток доксорубицина или озона активирует в злокачественных клетках печени процессы свободнорадикального окисления, что сопровождается увеличением продуктов липопероксидации. Их введение повышает концентрацию фермента каспазы-3 как при изолированном, так и при сочетанном действии, при этом в случае использования доксорубицина повышение количества каспазы-3 более существенно. Активация свободнорадикального окисления вызывает как некротические, так и апоптотические изменения в клетках культуры печени — через увеличение количества каспаз.
Финансирование исследования. Работа не финансировалась никакими источниками.
Литература
- Алясова А.В., Конторщикова К.Н., Терентьев И.Г., Иванова И.П., Кузнецов С.С., Сазанов А.И. Влияние низких терапевтических концентраций озонированного физиологического раствора на терапевтический патоморфоз опухоли в эксперименте. Современные технологии в медицине 2010; 4: 27–32.
- Красникова О.В., Гордецов А.С., Конторщикова К.Н., Крылов В.Н., Сазанов А.И. Изменение параметров ИК-спектров биологических тканей животных-опухоленосителей на фоне совместного введения доксорубицина и озона. Современные технологии в медицине 2011; 3: 83–87.
- Alyasova A.V., Vedunova M.V., Mishchenko T.A., Terentyev I.G., Tsybusov S.N., Kontorshchikova K.N. Effect of ozone and doxorubicin on the viability and morphology of malignant hepatic cells. Sovremennye tehnologii v medicine 2016; 8(2): 84–89, http://dx.doi.org/10.17691/stm2016.8.2.12.
- Конторщикова К.Н. Перекисное окисление липидов при коррекции гипоксических нарушений физико-химическими факторами. Автореф. дис. … докт. биол. наук. Н. Новгород; 1992.
- Масленников О.В., Конторщикова К.Н., Шахов Б.Е. Руководство по озонотерапии. Н. Новгород: Издательство “Исток”; 2015; 345 c.
- Конторщикова К.Н., Перетягин С.П. Закономерность формирования адаптационных механизмов организмов млекопитающих при системном воздействии низкими терапевтическими дозами озона. Диплом на открытие 309. 2006.
- Сарвилина И.В., Каркищенко В.Н., Горшкова Ю.В. Междисциплинарные исследования в медицине. М: Техносфера; 2007; c. 139–145.
- Кузьмина Е.И., Нелюбин А.С., Щенникова М.К. Применение индуцированной хемилюминесценции для оценок свободнорадикальных реакций в биологических субстратах. В кн.: Межвузовский сборник биохимии и биофизики микроорганизмов. Горький; 1983; c. 179–183.
- Волчегорский И.А., Налимов А.Г., Яровинский Б.Г., Лифшиц Р.И. Сопоставление различных подходов к определению продуктов ПОЛ в гептан-изопропанольных экстрактах крови. Вопросы медицинской химии 1989; 1: 127–131.








