Изменения сетевых тета- и гамма-осцилляций при развитии нейродегенеративных заболеваний (обзор)
В осуществлении когнитивных функций мозга участвует ритмическая тета- и гамма-активность, регистрируемая в неокортексе и гиппокампе; при многих нейрологических и психиатрических заболеваниях выявлены нарушения этой активности. Болезнь Альцгеймера (БА) и височная эпилепсия (ВЭ) являются наиболее распространенными формами нейродегенеративных заболеваний, характеризующимися прогрессивными нарушениями познавательных функций. Для успешного лечения этих заболеваний очень важна их ранняя диагностика. Проведено множество исследований для выявления признаков развития данных патологий, однако в настоящее время остается большая потребность в таких методах, которые выявляли бы изменения функционирования мозга в миллисекундных интервалах времени, чтобы получить информацию о специфических когнитивных нарушениях.
В настоящем обзоре рассматриваются литературные данные, показывающие, что когерентный анализ ритмической тета- и гамма-активности может быть использован при ранней диагностике БА и ВЭ. Эти данные демонстрируют, что в мозге с развивающимся эпилептическим очагом наблюдаются специфические нарушения когерентности тета- и гамма-осцилляций во многих нейронных сетях мозга: гиппокампальной, гиппокампально-энторинальной, гиппокампально-префронтальной и гиппокампально-септальной. На моделях БА обнаружены значительные изменения тета–гамма-когерентности в субикулуме — структуре, являющейся выходной структурой для гиппокампа; выявлены также нарушения когерентности тета- и гамма-осцилляций в гиппокампально-париетальной и гиппокампально-префронтальной сетях.
Данные, полученные посредством когерентного анализа, в совокупности с фактами, обнаруженными с помощью когнитивных тестов, можно использовать в ранней диагностике нейропатологий с дефицитом познавательных функций.
Введение
Известно, что тета- и гамма-осцилляции тесно связаны с когнитивными процессами. Тета-ритм (4–12 Гц) представляет собой высокоамплитудные осцилляции в гиппокампе и окружающих его лимбических структурах во время исследовательского поведения и парадоксального сна [1–3]. Тета-осцилляции регистрируются также в неокортексе, где они особенно отчетливы во фронтально-медиальных областях [4–8] и в субкортикальных структурах [9–13]. Тета-ритм участвует в формировании и извлечении эпизодической и пространственной памяти [14]. Гамма-осцилляции (30–100 Гц) обычно регистрируются одновременно с тета-ритмом в гиппокампе [15–17] и в неокортексе (чаще всего во фронтальных и париетальных областях) [18, 19]. Предполагают, что гамма-осцилляции играют роль в механизмах внимания [20, 21] и удержания информации в памяти во время выполнения когнитивных задач [22, 23]. Последние данные указывают на то, что связь между фазой медленных колебаний (в частности, тета) и амплитудой быстрых колебаний (гамма) может быть вовлечена в обработку информации [24–26].
Ключевым компонентом когнитивных процессов являются взаимодействия между структурами мозга [27–30]. Накапливающиеся данные указывают, что одним из фундаментальных механизмов коммуникации между структурами являются ритмы мозга. Это доказывается синхронизацией ритмических локальных полевых потенциалов (ЛПП) в различных областях мозга при выполнении задач, где необходимо участие внимания и памяти [31–37], и возрастанием при этом кросс-корреляции нейронных осцилляций [29, 30, 38]. Синхронизация ЛПП в разных областях мозга оказывает существенное влияние на способность этих областей взаимодействовать друг с другом [34]; объединяя активность пространственно разделенных, но функционально связанных нейронов, она также обеспечивает механизмы формирования клеточных ансамблей и их пространственно-временнóй координации [30, 39–42]. Кроме того, осцилляции могут рассматриваться как механизм ритмических изменений нейронной возбудимости [43, 44].
Гипотеза о коммуникации разных областей мозга посредством синхронизации [44] в настоящее время широко поддерживается [30, 36, 45–51]. По этой гипотезе анатомические связи могут быть эффективными или неэффективными в зависимости от наличия или отсутствия ритмической синхронизации [44, 52].
Известно, что взаимодействие структур мозга и осцилляторная активность в них могут нарушаться при психических и неврологических расстройствах [42, 53–57]. Однако, несмотря на десятилетия исследований, нарушения когерентности ритмов при таких патологиях, как болезнь Альцгеймера (БА) и височная эпилепсия (ВЭ), остаются неясными.
Когерентность тета- и гамма-ритмов
Кортикальные структуры генерируют большое количество осцилляций различной частоты. В то время как высокочастотные ритмы отражают локальную нейронную активность, в низкочастотных осцилляциях участвуют широко распределенные по мозгу области [42]. Внешние для организма или внутренние стимулы могут вызывать синхронизацию ритмов и таким образом приводить к формированию нового явления более высокого порядка, известного как фазовая сцепленность (phase coupling), или фазовая когерентность. Стандартная фазовая когерентность выявляет относительное постоянство фазовых различий между двумя осцилляциями одной и той же частоты, т.е. внутричастотную когерентность, или синхронность (рис. 1) [59, 60]. Показано, что фазовая когерентность отражает различные когнитивные процессы у людей, обезьян, крыс и мышей [25, 42, 61–66].
|
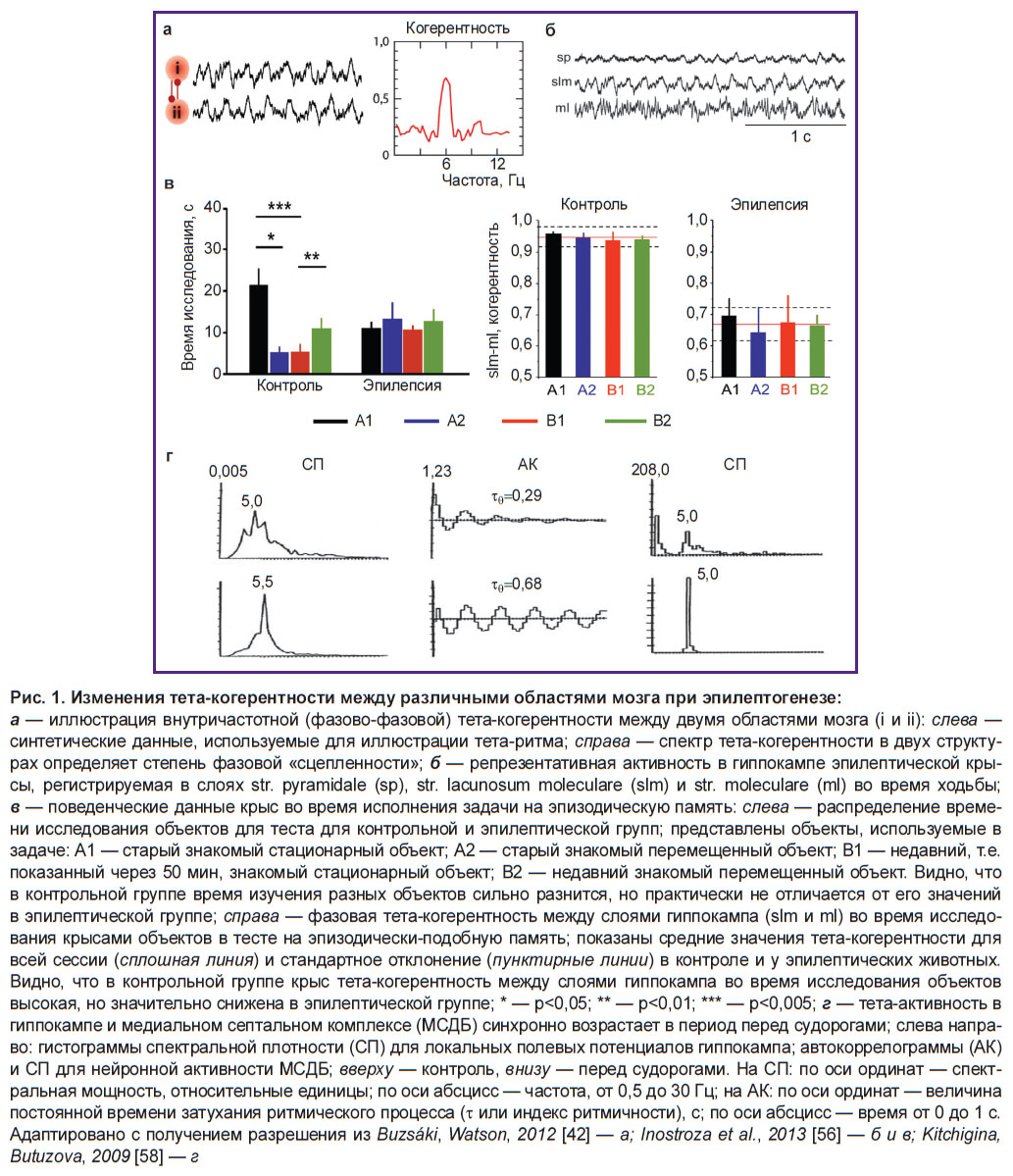
Рис. 1. Изменения тета-когерентности между различными областями мозга при эпилептогенезе: a — иллюстрация внутричастотной (фазово-фазовой) тета-когерентности между двумя областями мозга (i и ii): слева — синтетические данные, используемые для иллюстрации тета-ритма; справа — спектр тета-когерентности в двух структурах определяет степень фазовой «сцепленности»; б — репрезентативная активность в гиппокампе эпилептической крысы, регистрируемая в слоях str. pyramidale (sp), str. lacunosum moleculare (slm) и str. moleculare (ml) во время ходьбы; в — поведенческие данные крыс во время исполнения задачи на эпизодическую память: слева — распределение времени исследования объектов для теста для контрольной и эпилептической групп; представлены объекты, используемые в задаче: А1 — старый знакомый стационарный объект; А2 — старый знакомый перемещенный объект; В1 — недавний, т.е. показанный через 50 мин, знакомый стационарный объект; B2 — недавний знакомый перемещенный объект. Видно, что в контрольной группе время изучения разных объектов сильно разнится, но практически не отличается от его значений в эпилептической группе; справа — фазовая тета-когерентность между слоями гиппокампа (slm и ml) во время исследования крысами объектов в тесте на эпизодически-подобную память; показаны средние значения тета-когерентности для всей сессии (сплошная линия) и стандартное отклонение (пунктирные линии) в контроле и у эпилептических животных. Видно, что в контрольной группе крыс тета-когерентность между слоями гиппокампа во время исследования объектов высокая, но значительно снижена в эпилептической группе; * — р<0,05; ** — р<0,01; *** — р<0,005; г — тета-активность в гиппокампе и медиальном септальном комплексе (МСДБ) синхронно возрастает в период перед судорогами; слева направо: гистограммы спектральной плотности (СП) для локальных полевых потенциалов гиппокампа; автокоррелограммы (АК) и СП для нейронной активности МСДБ; вверху — контроль, внизу — перед судорогами. На СП: по оси ординат — спектральная мощность, относительные единицы; по оси абсцисс — частота, от 0,5 до 30 Гц; на АК: по оси ординат — величина постоянной времени затухания ритмического процесса (τ, или индекс ритмичности), с; по оси абсцисс — время от 0 до 1 с. Адаптировано с получением разрешения из Buzsáki, Watson, 2012 [42] — a; Inostroza et al., 2013 [56] — б и в; Kitchigina, Butuzova, 2009 [58] — г |
Внутричастотная когерентность между осцилляциями в различных областях мозга (рис. 1, a) интенсивно изучалась вследствие ее предполагаемой роли в регуляции межструктурных взаимодействий [34, 67–70].
Кроме этого, корреляция между амплитудными характеристиками двух волн на разной частоте, называемая кросс-частотной амплитудно-амплитудной когерентностью, является еще одной характеристикой осцилляций [71, 72]; этот тип взаимодействия описывался рядом авторов [71–74], но вопреки корреляции с поведением его функциональная роль остается малопонятной.
Фазовая когерентность между тета- и гамма-осцилляциями, а именно фазово-амплитудная кросс-частотная когерентность (КЧК) [7, 15, 22, 25, 64, 66, 75–83] и фазово-фазовая КЧК, в которой несколько гамма-циклов вовлечены в один тета-цикл [84–88], является наиболее изученным феноменом фазовой когерентности. Фазово-амплитудная КЧК описывает зависимость между фазой низкочастотного ритма и амплитудой высокочастотных осцилляций [24] (рис. 2). Таким образом, она отражает взаимодействия между локальной (микро) сетью [49, 90] и системной (макро) сетью [24, 78, 91].
|
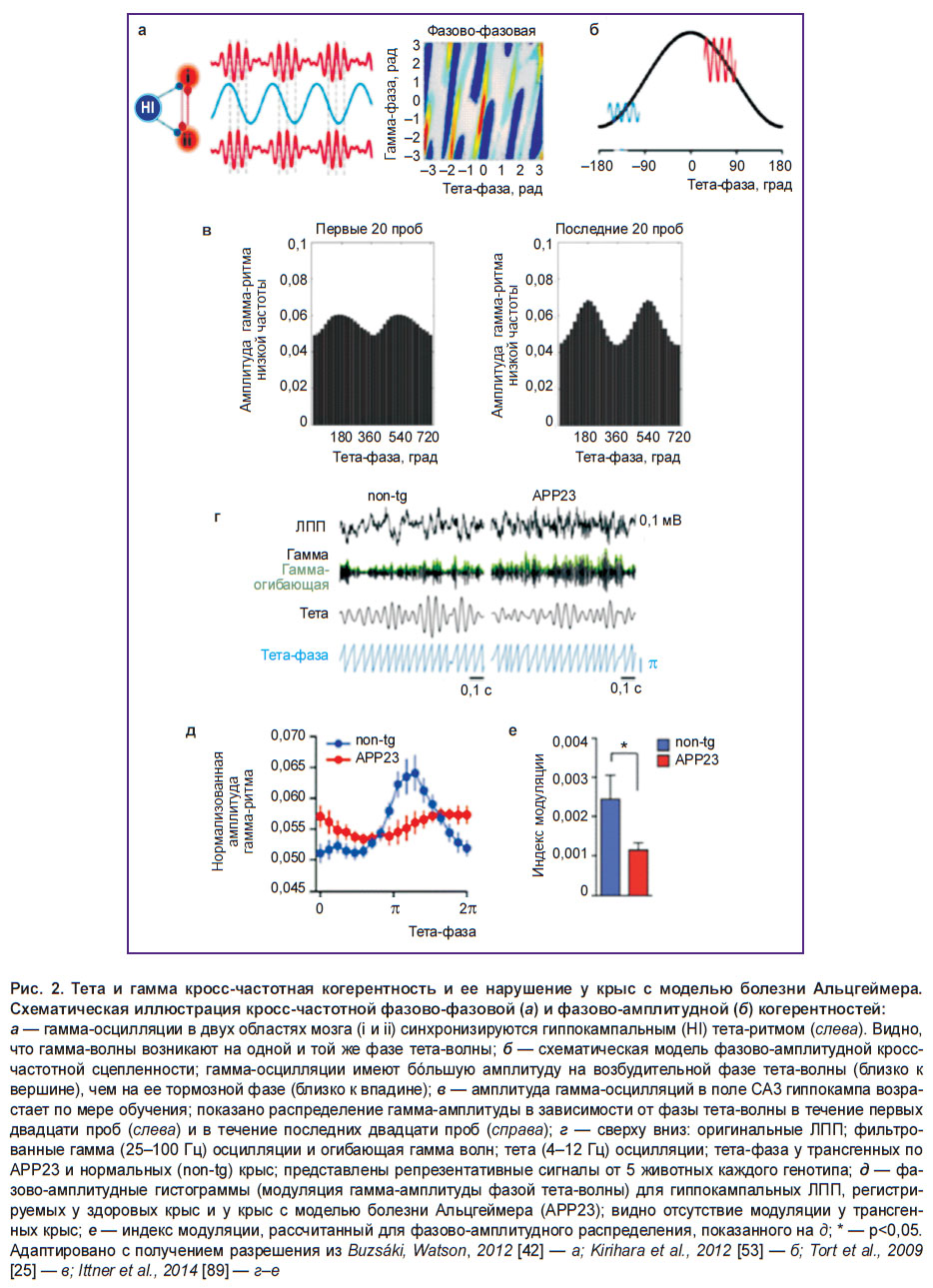
Рис. 2. Тета и гамма кросс-частотная когерентность и ее нарушение у крыс с моделью болезни Альцгеймера. Схематическая иллюстрация кросс-частотной фазово-фазовой (а) и фазово-амплитудной (б) когерентностей: а — гамма-осцилляции в двух областях мозга (i и ii) синхронизируются гиппокампальным (HI) тета-ритмом (слева). Видно, что гамма-волны возникают на одной и той же фазе тета-волны; б — схематическая модель фазово-амплитудной кросс-частотной сцепленности; гамма-осцилляции имеют бóльшую амплитуду на возбудительной фазе тета-волны (близко к вершине), чем на ее тормозной фазе (близко к впадине); в — амплитуда гамма-осцилляций в поле СА3 гиппокампа возрастает по мере обучения; показано распределение гамма-амплитуды в зависимости от фазы тета-волны в течение первых двадцати проб (слева) и в течение последних двадцати проб (справа); г — сверху вниз: оригинальные ЛПП; фильтрованные гамма (25–100 Гц) осцилляции и огибающая гамма волн; тета (4–12 Гц) осцилляции; тета-фаза у трансгенных по АРР23 и нормальных (non-tg) крыс; представлены репрезентативные сигналы от 5 животных каждого генотипа; д — фазово-амплитудные гистограммы (модуляция гамма-амплитуды фазой тета-волны) для гиппокампальных ЛПП, регистрируемых у здоровых крыс и у крыс с моделью болезни Альцгеймера (APP23); видно отсутствие модуляции у трансгенных крыс; е — индекс модуляции, рассчитанный для фазово-амплитудного распределения, показанного на д; * — р<0,05. Адаптировано с получением разрешения из Buzsáki, Watson, 2012 [42] — a; Kirihara et al.,2012 [53] — б; Tort et al.,2009 [25] — в; Ittner et al., 2014 [89] — г–е |
Это явление представляет собой наиболее отчетливый «закон», лежащий в основе иерархии в системе мозговых осцилляторов, где фаза более медленных осцилляций модулирует амплитуду более быстрого ритма (или ритмов) [15, 92, 93]. Можно констатировать, что фазово-амплитудная КЧК может использоваться как индекс кортикальной возбудимости и взаимодействия сетей [94–96]. В неэпилептогенном гиппокампе пациентов, подлежащих нейрохирургической операции, и в здоровом мозге грызунов степень тета и гамма фазово-амплитудной когерентности повышалась по мере их обучения [25, 64, 97]. Обычно в нормальном гиппокампе гамма- и тета-осцилляции демонстрируют значительную фазово-амплитудную КЧК, которая рассматривается как необходимое звено в осуществлении гиппокампальных функций [25, 64, 98]. Так, во время пространственного обучения выраженность гиппокампальной тета–гамма-когерентности обычно коррелирует с возрастанием правильности выполнения когнитивной задачи [25].
В то же время фазово-фазовая КЧК обеспечивает, как предполагается, физиологический механизм взаимодействий активности, генерируемой на существенно различающейся частоте. Так как гамма-осцилляции значительно быстрее, чем тета-осцилляции, то во время одного тета-цикла могут генерироваться множество гамма-циклов (рис. 2, a, б). Явление фазово-фазовой тета–гамма-сцепленности означает, что гамма-волны всегда возникают во время одной и той же фазы тета-волн. Предполагается также, что фазово-фазовая КЧК принимает участие в таких когнитивных процессах, как внимание и память [22, 71, 78, 99, 100]. Более десяти лет назад J.E. Lisman и M.A. Idiart была представлена интересная нейронная модель, в которой емкость рабочей памяти («7±2») определялась количеством гамма-циклов в одном тета-цикле («n:m») [78]. Самые последние исследования указывают на то, что гиппокамп действительно использует механизм кодирования информации посредством фазово-фазовой тета–гамма-когерентности [85–88]. Констатируется, что временнáя координация нейронных спайков фазово-фазовой тета–гамма-сцепленностью может облегчать передачу информации и спайк-тайминг-зависимую пластичность [100–102]. В то же время при некоторых нейродегенеративных патологиях может наблюдаться десинхронизация ритмов.
Нарушения когерентности тета- и гамма-ритмов, типичные для болезни Альцгеймера и височной эпилепсии
Изменения когерентности ритмов в эпилептическом мозге. Эпилепсия является заболеванием, характеризующимся возрастанием нейронной возбудимости, клеточной гибелью и образованием аберрантных связей в мозге (см. обзор [103]). Наиболее распространенной и фармакологически резистентной формой фокальной эпилепсии у взрослых считается височная эпилепсия (ВЭ). У пациентов с ВЭ выявлено селективное и значительное нарушение эпизодической (автобиографической) памяти, для которой характерно то, что запоминаемые события во время кодирования и извлечения информации располагаются во временнóм контексте [104]. Так же, как и люди, эпилептические животные в значительной степени обнаруживают селективное повреждение эпизодически-подобной памяти при относительной сохранности у них других форм гиппокамп-зависимой памяти [105–107].
Гиппокампальная сеть. Анализ ЛПП, зарегистрированных в гиппокампе у здоровых пациентов во время исполнения тестов на эпизодическую память, выявил резкое повышение гамма-осцилляций перед успешным запоминанием событий. В то же время в гиппокампе пациентов с ВЭ, напротив, в этом случае наблюдалось снижение мощности гамма-ритма [97, 108]. Таким образом, при исполнении тестов на эпизодическую память у людей с ВЭ обнаружена реверсия типичных изменений гиппокампальных гамма-осцилляций [108]. Кроме того, было показано на каиновой модели [56], что у крыс с дефицитом эпизодически-подобной памяти, вызванным введением каината (что провоцирует развитие эпилептического очага), в гиппокампе наблюдается редукция мощности тета-ритма и тета-когерентности вдоль оси «поле CA1–зубчатая фасция». Конкретнее, у эпилептических животных наблюдалось снижение когерентности тета-осцилляций между слоями str. lacunosum-moleculare (slm) поля CА1 и str. moleculare (ml) зубчатой фасции (ЗФ) по сравнению с контрольными крысами (рис. 1, б, в). М. Inostroza с соавт. [56] полагают, что, поскольку в slm и ml заканчиваются прямые входы от III и IIслоев энторинальной коры (ЭК) и от контралатерального гиппокампа, то дискоординация этих входов у эпилептических животных — наиболее вероятная причина нарушения у них эпизодической памяти.
Гиппокампально-энторинальная сеть. Известно, что ключевым механизмом эпизодической памяти является синхронизация нейрональной активности в сети гиппокамп–ЭК; именно этот механизм повреждается при ВЭ [105]. У свободно-подвижных эпилептических животных с каиновой моделью ВЭ выявлено изменение тета-когерентности между ЭК и ЗФ во время интериктальной фазы [54]. А именно, у эпилептических мышей тета-активность в ЭК отставала по фазе от активности в ЗФ; в то же время у здоровых животных активность между ЭК и ЗФ была синхронной, что проявлялось в отчетливом внутричастотном фазовом сцеплении. Используя компьютерную модель, авторы предположили, что гибель гиппокампальных нейронов нарушает взаимодействия гиппокампальных подсетей, что вызывает ЭК–ЗФ-дискоординацию [54].
В здоровом мозге входы от медиальной и латеральной ЭК через темпороаммонов и перфорирующий пути вызывают разряды гиппокампальных нейронов, которые отражают интегрированное представительство пространственной и временнóй информации [109–113], а также недавний опыт, приобретенный животным или человеком [114, 115]. Это нейронное кодирование точно организовано во временнóй шкале, что осуществляется текущими осцилляциями, особенно тета- и гамма-ритмами [2, 14, 26, 116–119]. Тщательное измерение проксимально-дистальной когерентности тета-активности в дорзальном гиппокампе здоровых и эпилептических крыс выявило, что у нормальных животных более сильная синхронизация между темпороаммоновым и перфорирующим входами наблюдается возле поля СА3 (в проксимальных локусах), в то время как у эпилептических — возле субикулума (в дистальных локусах) [57]. Этот противоположный тренд у крыс с патологией ассоциировался с реорганизацией темпороаммонова и перфорирующего путей, которая сопровождала выявленную впоследствии гибель гиппокампальных клеток. F. Laurent с соавт. [57] также продемонстрировали нарушение у эпилептических крыс временнóй координации сигналов, приходящих в гиппокамп от ЭК с задержкой в несколько тета-циклов на проксимальном и дистальном концах гиппокампа. Таким образом, авторы показали, что проксимально-дистальная организация разных входов в гиппокамп от ЭК играет важную роль в функционировании височной области мозга и что в эпилептическом мозге она повреждается.
Гиппокампально-префронтальная сеть. Как упоминалось выше, тета- и гамма-осцилляции в норме присутствуют в гиппокампе и медиальной префронтальной коре (мПФК) и синхронно изменяются при выполнении различных когнитивных задач [19, 64]. Гиппокампальные тета-волны обычно сцеплены с тета-волнами мПФК [19] и модулируют гамма-осцилляции в гиппокампе и префронтальной коре во время когнитивного поведения [64, 120, 121]. У животных с моделью ВЭ, провоцированной стимуляцией перфорирующего пути, наблюдались аномальные изменения в сети гиппокамп–мПФК при регистрации полевой активности у крыс со спонтанными судорогами [122]. Авторы выявили, что возвратные судороги ослабляют текущий гиппокампальный тета-ритм; в то же время непосредственно перед генерацией самих судорог тета-когерентность между гиппокампом и мПФК сильно возрастает. Одновременно с возрастанием тета-синхронности усиливается кросс-частотная сцепленность между тета-ритмом в гиппокампе и гамма-ритмом — в мПФК. Используя показатель причинности Грэнжера (Granger causality), авторам удалось показать, что увеличение синхронности осцилляций в гиппокампе и мПФК в преиктальной фазе провоцировалось гиппокампальной сетью [122]. Слишком сильная синхронность между тета-ритмом в гиппокампе и гамма-риммом в префронтальной коре могла быть причиной аномальной пластичности в мПФК-сетях, которая, как предполагается, в свою очередь вызывала изменения, наблюдаемые у нейронов этой области неокортекса [86].
Гиппокампально-септальная сеть. Регистрация ЛПП в гиппокампе и медиальном септальном комплексе (медиальное септальное ядро и ядро диагонального пучка Брока — МСДБ) выявила, что в этих областях здорового мозга тета-осцилляции обычно синхронны [10, 13, 36]. Как правило, амплитуда тета-осцилляций в МСДБ была ниже, чем в гиппокампе, но частота тета-ритма сильно не различалась. Тета-когерентность между гиппокампом и МСДБ была достаточно высокой: фазовый анализ активности у здоровых животных не выявил существенного (<10 мс) однонаправленного сдвига между фазами тета-волн в этих структурах [10, 13]. У хронических эпилептических животных значительное снижение мощности тета-ритма обнаружено в гиппокампе [36, 123–126] и МСДБ [13]. Кроме того, на пилокарпиновой модели ВЭ обнаружено снижение синхронности септальных и гиппокампальных ЛПП [127]. Однако на модели ВЭ, генерированной киндлингом перфорирующего пути, было показано повышение тета-синхронности между гиппокампом и МСДБ во время эпилептогенеза [13]. Более того, на этой модели ВЭ у кроликов наблюдалось драматическое повышение тета-осцилляций одновременно в гиппокампе и МСДБ непосредственно перед (не более чем за 20 с) возникновением судорожной активности [58]. Это явление напоминает события, показанные для гиппокампально-префронтальной сети перед началом судорог у животных с аналогичной моделью ВЭ [122]. Интересно, что на модели ВЭ, провоцированной у морской свинки киндлингом перфорирующего пути, взаимодействия между гиппокампом и МСДБ при эпилептогенезе изменялись на противоположные: в начале киндлинга МСДБ опережала гиппокамп по фазе тета-волны, а после формирования патологического очага отставала от него [13]. В дополнение к этому взаимоотношения между ритмическими залпами септальных нейронов и фазами гиппокампальных тета-волн во время спонтанных судорог изменялись на противоположные по сравнению с интериктальным периодом [58]; т.е. эти отношения не были постоянными.
Нарушение когерентности ритмов при болезни Альцгеймера: клинические и экспериментальные данные
Изменения тета- и гамма-ритмов в мозге при болезни Альцгеймера. БА представляет собой прогрессирующее нейродегенеративное заболевание, при котором наблюдаются необратимые нарушения когнитивных функций, особенно памяти. Этиология БА неизвестна, и в настоящее время в медицине отсутствуют надежные способы лечения данного заболевания. В начале этого века появилась так называемая амилоидная гипотеза [128, 129], в которой предполагается, что бета-амилоидный (Aβ) пептид, главный компонент амилоидных бляшек, ответственен за нарушения когнитивных функций при БА [129, 130]. Однако в последние годы этой гипотезе были противопоставлены факты, свидетельствующие о том, что в основе этиологии БА могут лежать нарушения метаболизма белка АРР (amyloid precursor protein), являющегося предшественником бета-амилоида и концентрирующегося в нейронных синапсах, а также прогрессирование этих нарушений за счет параллельно развивающейся патологии тау-белка (см. обзор [131]). Более того, недавно было обнаружено, что амилоидные бляшки проявляют способность связывать вновь образующиеся бета-амилоиды [132], т.е. обладают протекторным свойством.
При БА нарушаются различные формы памяти [133]. Однако это поисходит и при других нейродегенеративных заболеваниях. Предполагается, что выраженный пространственный дефицит в некоторой степени может помочь идентифицировать пациентов с большим риском развития БА и отличить их от субъектов с другими болезнями, при которых также нарушается память [134]. В некоторых работах показано, что у пациентов с БА в отличие от субъектов с нормальным старением выявляется повышение относительной мощности медленных осцилляций (в частности, тета-ритма) и снижение относительной мощности быстрых осцилляций (гамма-ритма) [135–139]. В других работах, напротив, у пациентов с БА обнаружено повышение мощности гамма-ритма и отсутствие повышения тета-осцилляций [140, 141]. Некоторые авторы отмечали, что изменения в ЭЭГ пациентов с различными видами деменции также могут характеризоваться подобными сетевыми нарушениями [136]. Кроме того, в противоположность данным, полученным на пациентах, у трансгенных (Tg5xFAD) мышей с моделью БА выявлено снижение выраженности как тета-, так и гамма-ритмов; в этом случае снижение мощности осцилляций предшествовало падению способности к обучению пространственным задачам [142]. Более того, у трансгенных (APP23) мышей с другой моделью БА продемонстрированы изменения ритмов (по сравнению с контролем), противоположные таковым у пациентов с БА: значительное снижение мощности тета-ритма (имеющего частоту ~10 Гц) и повышение мощности гамма-осцилляций (с частотой 25–50 Гц) [93]. Таким образом, снижение или повышение выраженности тета- и гамма-осцилляций как таковых может быть неспецифичным для этой патологии [136].
Изменения тета–гамма-когерентности в мозге с патологией, характерной для болезни Альцгеймера. Можно предположить, что наиболее убедительными свидетельствами нарушений ритмов в мозге при БА являются изменения тета–гамма-КЧК. Так, у пациентов с БА кроме возрастания гамма-ритма в ЭЭГ выявлено повышение кросс-частотной сцепленности между активностью в гамма-полосе и низкочастотной (в частности, тета) активностью по сравнению со здоровыми добровольцами [141]. Во время выполнения задачи на рабочую память были получены свидетельства связи между нарушением тета–гамма-когерентности и дефицитом рабочей памяти у пациентов с БА [143].
Кроме того, у взрослых трансгенных (APP23) свободно-перемещающихся мышей наблюдалось нарушение модуляции гамма-осцилляций гиппокампальным тета-ритмом (рис. 2, в, г) [89]. Важно, что эти изменения наблюдались до образования амилоидных бляшек. Более того, у значительной части трансгенных (TgCRND8) мышей с другой моделью БА в месячном возрасте обнаружено существенное изменение тета–гамма-сцепленности в субикулуме [144]. Такое «разъединение» выявлялось до проявления каких-либо гистологических нарушений, в частности образования амилоидных бляшек. Кроме того, у используемых в экспериментах молодых мышей (в возрасте 1 мес) экспрессируется чрезвычайно низкий уровень бета-амилоида. R. Goutagny с соавт. [144] полагают, что нарушение тета–гамма-КЧК в субикулуме может рассматриваться как наиболее ранний биомаркер развивающейся БА. Такое предположение противоречит существующей (упоминавшейся выше) гипотезе, где заявлялось, что дефицит памяти и сетевые нарушения на моделях БА у животных вызываются гиперпродукцией растворимого бета-амилоида [129, 130, 145].
Интересно, что хотя белок АРР, как считается в настоящее время, вовлечен в патофизиологию БА, дефицит его у мышей вызывает нарушения когнитивных функций [146, 147]. Это подтверждает, что в здоровом мозге АРР играет существенную роль в работе нейронных сетей. В недавнем исследовании у нокаутных по АРР мышей была обнаружена существенно сниженная тета–гамма-сцепленность ЛПП, регистрируемых в дорзальном гиппокампе и париетальной коре; кроме того, у этих нокаутных мышей была значительно нарушена гиппокампально-префронтальная КЧК [148]. Эти изменения могут играть роль в возникновении когнитивных нарушений у животных с дефицитом АРР. Таким образом, APP важен для взаимодействия ритмов, генерируемых в различных полосах частот. Факты, изложенные выше, могут означать, что между функционированием АРР в здоровом мозге и мозге с БА-патологией существует лишь очень тонкая граница.
В недавней работе [73] было протестировано, является ли агрегация гиперфосфорилированного тау-белка в энторинальной коре (как преклинического признака БА) причиной нарушения координации ЛПП между эфферентными для этой области структурами — гиппокампом и прелимбической мПФК. У здоровых животных выявлено усиление фазово-фазовой сцепленности тета–гамма-осцилляций в этих двух областях во время ассоциативного обучения (крысы обучались ассоциировать два стимула, разделенных коротким временным интервалом). В отличие от нормальных крыс у тау-экспрессирующих животных в гиппокампе и мПФК обнаружено значительное ослабление вызванного стимулом тета-ритма. При этом несмотря на сохранение нормального запоминания, связанная с обучением сцепленность осцилляций между гиппокампом и мПФК снижалась; в то же время в пределах мПФК усиливалась тета–гамма-фазово-амплитудная когерентность. Авторы полагают, что тау-агрегация в ЭК вызывает во время обучения аберрантную активность в обширных нейронных сетях, в которые вовлечена эта структура [73]. Результаты, полученные в данной работе, указывают на нарушения нейронных осцилляций в преклинической стадии развития БА, что может приводить к прогрессированию заболевания.
Сходства и различия нарушений в мозге с нейропатологиями, характерными для болезни Альцгеймера и височной эпилепсии
Сходство в нарушениях осцилляторной активности в мозге с БА и ВЭ (в частности, нарушения тета–гамма-когерентности в гиппокампально-кортикальных сетях) позволяет предполагать, что эти заболевания имеют некоторые общие свойства.
Потенциальная связь между ВЭ и БА подтверждается экспериментальными и клиническими данными. Так, старение является общим и хорошо установленным фактором риска для эпилепсии и БА [149–153]. Кроме того, БА может быть важной причиной развития эпилепсии, что показано у пожилых людей [149, 154–156] и у животных с моделью БА [153, 157]. У пациентов с БА фактор риска для развития судорог или других форм эпилептиформной активности в 5–10 раз выше [150]. Хотя судорожная патология ранее рассматривалась как вторичная по отношению к БА, впоследствии было выявлено, что активность нейронов регулирует их чувствительность к бета-амилоидам [129, 158, 159]; более того, повышенная нейронная возбудимость увеличивает генерацию Aβ [160]. Эпилептиформная активность может способствовать развитию когнитивных нарушений: в сочетании с нарушенными ритмами в височной области (особенно в гиппокампе) она способна вызывать амнезию, что ослабляется антиэпилептическими препаратами [55, 161]. У пациентов с судорогами и БА были описаны случаи с феноменом “vu/déjà vu” (уже виденного) [162]; в другой работе [163] показано явление, называемое «эпилептической продромальной БА». Авторы полагают, что это эпилептическая разновидность БА, которая обычно начинается с судорог, а затем сопровождается когнитивным дефицитом. Сходные признаки когнитивных и поведенческих нарушений при ВЭ и БА приведены в недавней работе [164].
Накоплено большое количество клинических данных, которые указывают на возможность возникновения ВЭ как сопутствующего заболевания у пациентов с БА; становится все более явным, что БА ассоциируется с нейрональной гипервозбудимостью, являющейся одной из основных причин развития эпилепсии [165–168]. Действительно, пациенты с эпилептической продромальной БА иногда страдают от судорог до того, как развиваются отчетливые когнитивные нарушения. Эпилептиформная активность может проявляться на ранних стадиях развития БА чаще, чем это предполагалось. Так, судороги у пациентов с БА и малыми амнестическими расстройствами ассоциируются с ранним проявлением ослабления когнитивных функций [150, 162, 169, 170]. В то же время нейродегенеративные процессы, свойственные деменции, могут играть центральную роль в развитии эпилепсии у пациентов, предрасположенных к когнитивному дефициту. Таким образом, возникающая у взрослых эпилепсия неизвестного происхождения может представлять собой фактор риска для развития нейродегенерации, даже если эпилептические проявления и клинически распознаваемая деменция разделены длительным промежутком времени [171].
В гиппокампе, одном из основных локусов гибели нейронов в мозге с БА или ВЭ, сетевая гиперсинхронизация и эпилептиформная активность могут быть результатом формирования обширных аберрантных межнейронных связей. Такое патологическое ремоделирование было обнаружено как у эпилептических крыс, так и у трансгенных по APP мышей [39, 129, 158, 172, 173]. Аберрантная реконструкция может быть причиной изменений осцилляторной активности и когерентности ритмов мозга у пациентов с ВЭ и БА и характерными для них патологиями.
Заключение
Известно, что основная проблема диагностики нейродегенеративных заболеваний — выявление нейронных повреждений на начальных стадиях их развития. К настоящему времени имеются определенные достижения в использовании таких методов ранней диагностики ВЭ и БА, как измерения цереброспинальной жидкости и плазмы, а также позитронно-эмиссионная томография. Однако остается большая потребность в биомаркерах, которые отражали бы изменения в функционировании мозга в течение очень коротких интервалов времени, таких как миллисекунды, чтобы получить информацию о прогрессирующем когнитивном дефиците [174]. Применение магнитоэнцефалографии [175] в комбинации с визуализацией мозга, особенно во время когнитивных нагрузок, может быть перспективным подходом для ранней диагностики этих заболеваний. Так, при эпилептизации мозга выявлены специфические нарушения взаимодействий тета- и гамма-ритмов в гиппокампальной, гиппокампально-энторинальной, гиппокампально-префронтальной и гиппокампально-септальной сетях. На модели БА обнаружено существенное изменение тета–гамма-сцепленности в субикулуме — структуре, являющейся выходной для гиппокампа; кроме того, показана сниженная тета–гамма-когерентность между гиппокампом и париетальной корой, а также между гиппокампом и префронтальной корой.
Можно думать, что в будущем специфические нарушения когерентности будут служить четким маркером повреждения селективных групп нейронов в мозге и позволят направлять терапевтические воздействия строго на определенные нейронные локусы для предотвращения развития болезни, избегая в то же время возникновения побочных эффектов. Кроме того, исследования нарушений когерентных осцилляций будут способствовать более глубокому пониманию механизмов развития БА и ВЭ и механизмов функционирования мозга в целом.
Финансирование исследования. Работа была поддержана грантом Российского фонда фундаментальных исследований (№18-015-00157-а) и совместным грантом Российского фонда фундаментальных исследований и Министерства инноваций и инвестиций Московской области (№17-44-500312 р_а).
Конфликт интересов отсутствует.
Литература
- Vanderwolf C. Hippocampal electrical activity and voluntary movement in the rat. Electroencephalogr Clin Neurophysiol 1969; 26(4): 407–418, https://doi.org/10.1016/0013-4694(69)90092-3.
- Bland B.H. The physiology and pharmacology of hippocampal formation theta rhythms. Prog Neurobiol 1986; 26(1): 1–54, https://doi.org/10.1016/0301-0082(86)90019-5.
- Buzsáki G. Theta oscillations in the hippocampus. Neuron 2002; 33(3): 325–340, https://doi.org/10.1016/s0896-6273(02)00586-x.
- Klimesch W., Doppelmayr M., Schimke H., Ripper B. Theta synchronization and alpha desynchronization in a memory task. Psychophysiology 1997; 34(2): 169–176, https://doi.org/10.1111/j.1469-8986.1997.tb02128.x.
- Kahana M.J., Sekuler R., Caplan J.B., Kirschen M., Madsen J.R. Human theta oscillations exhibit task dependence during virtual maze navigation. Nature 1999; 399(6738): 781–784, https://doi.org/10.1038/21645.
- Jensen O., Tesche C.D. Frontal theta activity in humans increases with memory load in a working memory task. Eur J Neurosci 2002; 15(8): 1395–1399, https://doi.org/10.1046/j.1460-9568.2002.01975.x.
- Vinogradova O.S. Expression, control, and probable functional significance of the neuronal theta-rhythm. Prog Neurobiol 1995; 45(6): 523–583, https://doi.org/10.1016/0301-0082(94)00051-i.
- Guderian S., Schott B.H., Richardson-Klavehn A., Duzel E. Medial temporal theta state before an event predicts episodic encoding success in humans. Proc Natl Acad Sci U S A 2009; 106(13): 5365–5370, https://doi.org/10.1073/pnas.0900289106.
- Paré D. Amygdala oscillations and the consolidation of emotional memories. Trends Cogn Sci 2002; 6(7): 306–314, https://doi.org/10.1016/s1364-6613(02)01924-1.
- Nerad L., McNaughton N. The septal EEG suggests a distributed organization of the pacemaker of hippocampal theta in the rat. Eur J Neurosci 2006; 24(1): 155–166, https://doi.org/10.1111/j.1460-9568.2006.04902.x.
- Magill P.J., Sharott A., Bolam J.P., Brown P. Delayed synchronization of activity in cortex and subthalamic nucleus following cortical stimulation in the rat. J Physiol 2006; 574(3): 929–946, https://doi.org/10.1113/jphysiol.2006.110379.
- DeCoteau W.E., Thorn C., Gibson D.J., Courtemanche R., Mitra P., Kubota Y., Graybiel A.M. Learning-related coordination of striatal and hippocampal theta rhythms during acquisition of a procedural maze task. Proc Natl Acad Sci U S A 2007; 104(13): 5644–5649, https://doi.org/10.1073/pnas.0700818104.
- Kabanova I.V., Sinelnikova V.V., Popova I.Y., Kichigina V.F., Aliev R.R. Coherence and phase analysis of theta oscillations in the septohippocampal system during generation of convulsive activity. Neurosci Behav Physi 2013; 43(2): 214–218, https://doi.org/10.1007/s11055-013-9716-1.
- Hasselmo M.E., Bodelón C., Wyble B.P. A proposed function for hippocampal theta rhythm: separate phases of encoding and retrieval enhance reversal of prior learning. Neural Comput 2002; 14(4): 793–817, https://doi.org/10.1162/089976602317318965.
- Bragin A., Jando G., Nadasdy Z., Hetke J., Wise K., Buzsaki G. Gamma (40–100 Hz) oscillation in the hippocampus of the behaving rat. J Neurosci 1995; 15(1): 47–60, https://doi.org/10.1523/jneurosci.15-01-00047.1995.
- Strogatz S.H. Nonlinear dynamics and chaos: with applications to physics, biology, chemistry and engineering. Cambridge, MA: Perseus Books; 2003.
- Buzsáki G., Wang X.-J. Mechanisms of gamma oscillations. Annu Rev Neurosci 2012; 35(1): 203–225, https://doi.org/10.1146/annurev-neuro-062111-150444.
- Bouyer J., Montaron M., Rougeul A. Fast fronto-parietal rhythms during combined focused attentive behaviour and immobility in cat: cortical and thalamic localizations. Electroencephalogr Clin Neurophysiol 1981; 51(3): 244–252, https://doi.org/10.1016/0013-4694(81)90138-3.
- Benchenane K., Peyrache A., Khamassi M., Tierney P.L., Gioanni Y., Battaglia F.P., Wiener S.I. Coherent theta oscillations and reorganization of spike timing in the hippocampal-prefrontal network upon learning. Neuron 2010; 66(6): 921–936, https://doi.org/10.1016/j.neuron.2010.05.013.
- Fries P. Neuronal gamma-band synchronization as a fundamental process in cortical computation. Annu Rev Neurosci 2009; 32(1): 209–224, https://doi.org/10.1146/annurev.neuro.051508.135603.
- Jutras M.J., Fries P., Buffalo E.A. Gamma-band synchronization in the macaque hippocampus and memory formation. J Neurosci 2009; 29(40): 12521–12531, https://doi.org/10.1523/jneurosci.0640-09.2009.
- Sauseng P., Klimesch W., Heise K.F., Gruber W.R., Holz E., Karim A.A., Glennon M., Gerloff C., Birbaumer N., Hummel F.C. Brain oscillatory substrates of visual short-term memory capacity. Curr Biol 2009; 19(21): 1846–1852, https://doi.org/10.1016/j.cub.2009.08.062.
- Sridharan D., Knudsen E.I. Gamma oscillations in the midbrain spatial attention network: linking circuits to function. Curr Opin Neurobiol 2015; 31: 189–198, https://doi.org/10.1016/j.conb.2014.11.006.
- Canolty R.T., Knight R.T. The functional role of cross-frequency coupling. Trends Cogn Sci 2010; 14(11): 506–515, https://doi.org/10.1016/j.tics.2010.09.001.
- Tort A.B.L., Komorowski R.W., Manns J.R., Kopell N.J., Eichenbaum H. Theta–gamma coupling increases during the learning of item-context associations. Proc Natl Acad Sci 2009; 106(49): 20942–20947, https://doi.org/10.1073/pnas.0911331106.
- Lisman J.E., Jensen O. The theta–gamma neural code. Neuron 2013; 77(6): 1002–1016, https://doi.org/10.1016/j.neuron.2013.03.007.
- Livanov M.N., Krylov V.Yu., Ostrjakova T.V., Shulgina G.I. Slow field potential oscillations as one of the basic mechanisms of integrative activity of neurons [proceedings]. Act Nerv Super (Praha) 1977; 19(1): 43–44.
- Vinogradova O.S. Hippocampus as comparator: role of the two input and two output systems of the hippocampus in selection and registration of information. Hippocampus 2001; 11(5): 578–598, https://doi.org/10.1002/hipo.1073.abs.
- Engel A.K., Fries P., Singer W. Dynamic predictions: oscillations and synchrony in top–down processing. Nat Rev Neurosci 2001; 2(10): 704–716, https://doi.org/10.1038/35094565.
- Igarashi K.M., Lu L., Colgin L.L., Moser M.-B., Moser E.I. Coordination of entorhinal–hippocampal ensemble activity during associative learning. Nature 2014; 510(7503): 143–147, https://doi.org/10.1038/nature13162.
- Fell J., Klaver P., Lehnertz K., Grunwald T., Schaller C., Elger C.E., Fernández G. Human memory formation is accompanied by rhinal–hippocampal coupling and decoupling. Nat Neurosci 2001; 4(12): 1259–1264, https://doi.org/10.1038/nn759.
- Varela F., Lachaux J.-P., Rodriguez E., Martinerie J. The brainweb: phase synchronization and large-scale integration. Nat Rev Neurosci 2001; 2(4): 229–239, https://doi.org/10.1038/35067550.
- Buzsáki G. Large-scale recording of neuronal ensembles. Nat Neurosci 2004; 7(5): 446–451, https://doi.org/10.1038/nn1233.
- Womelsdorf T., Fries P., Mitra P.P., Desimone R. Gamma-band synchronization in visual cortex predicts speed of change detection. Nature 2005; 439(7077): 733–736, https://doi.org/10.1038/nature04258.
- Vinck M., Bos J.J., Van Mourik-Donga L.A., Oplaat K.T., Klein G.A., Jackson J.C., Gentet L.J., Pennartz C.M. Cell-type and state-dependent synchronization among rodent somatosensory, visual, perirhinal cortex, and hippocampus CA1. Front Syst Neurosci 2016; 9: 187, https://doi.org/10.3389/fnsys.2015.00187.
- Astasheva E.V., Kitchigina V.F. Activation of the glutamatergic system of the medial septal area accelerates epileptogenesis. Neurosci Behav Physi 2011; 41(3): 316–320, https://doi.org/10.1007/s11055-011-9418-5.
- Bott J.B., Muller M.A., Jackson J., Aubert J., Cassel J.C., Mathis C., Goutagny R. Spatial reference memory is associated with modulation of theta–gamma coupling in the dentate gyrus. Cereb Cortex 2015; 26(9): 3744–3753, https://doi.org/10.1093/cercor/bhv177.
- Tabuchi E.T., Mulder A.B., Wiener S.I. Position and behavioral modulation of synchronization of hippocampal and accumbens neuronal discharges in freely moving rats. Hippocampus 2000; 10(6): 717–728, https://doi.org/10.1002/1098-1063(2000)10:6717::aid-hipo10093.0.co;2-3.
- Harris K.D., Csicsvari J., Hirase H., Dragoi G., Buzsáki G. Organization of cell assemblies in the hippocampus. Nature 2003; 424(6948): 552–556, https://doi.org/10.1038/nature01834.
- Colgin L.L., Moser E.I. Gamma oscillations in the hippocampus. Physiology 2010; 25(5): 319–329, https://doi.org/10.1152/physiol.00021.2010.
- Buzsáki G. Neural syntax: cell assemblies, synapsembles, and readers. Neuron 2010; 68(3): 362–385, https://doi.org/10.1016/j.neuron.2010.09.023.
- Buzsáki G., Watson B.O. Brain rhythms and neural syntax: implications for efficient coding of cognitive content and neuropsychiatric disease. Dialogues Clin Neurosci 2012; 14(4): 345–367.
- Volgushev M., Chistiakova M., Singer W. Modification of discharge patterns of neocortical neurons by induced oscillations of the membrane potential. Neuroscience 1998; 83(1): 15–25, https://doi.org/10.1016/s0306-4522(97)00380-1.
- Fries P. A mechanism for cognitive dynamics: neuronal communication through neuronal coherence. Trends Cogn Sci 2005; 9(10): 474–480, https://doi.org/10.1016/j.tics.2005.08.011.
- Jensen O., Kaiser J., Lachaux J.-P. Human gamma-frequency oscillations associated with attention and memory. Trends Neurosci 2007; 30(7): 317–324, https://doi.org/10.1016/j.tins.2007.05.001.
- Mitchell D.J., McNaughton N., Flanagan D., Kirk I.J. Frontal-midline theta from the perspective of hippocampal “theta”. Prog Neurobiol 2008; 86(3): 156–185, https://doi.org/10.1016/j.pneurobio.2008.09.005.
- Rutishauser U., Ross I.B., Mamelak A.N., Schuman E.M. Human memory strength is predicted by theta-frequency phase-locking of single neurons. Nature 2010; 464(7290): 903–907, https://doi.org/10.1038/nature08860.
- Wang X.-J. Neurophysiological and computational principles of cortical rhythms in cognition. Physiol Rev 2010; 90(3): 1195–1268, https://doi.org/10.1152/physrev.00035.2008.
- Colgin L.L., Denninger T., Fyhn M., Hafting T., Bonnevie T., Jensen O., Moser M.B., Moser E.I. Frequency of gamma oscillations routes flow of information in the hippocampus. Nature 2009; 462(7271): 353–357, https://doi.org/10.1038/nature08573.
- Colgin L.L. Oscillations and hippocampal–prefrontal synchrony. Curr Opin Neurobiol 2011; 21(3): 467–474, https://doi.org/10.1016/j.conb.2011.04.006.
- Colgin L.L. Theta–gamma coupling in the entorhinal–hippocampal system. Curr Opin Neurobiol 2015; 31: 45–50, https://doi.org/10.1016/j.conb.2014.08.001.
- Bastos A.M., Vezoli J., Fries P. Communication through coherence with inter-areal delays. Curr Opin Neurobiol 2015; 31: 173–180, https://doi.org/10.1016/j.conb.2014.11.001.
- Kirihara K., Rissling A.J., Swerdlow N.R., Braff D.L., Light G.A. Hierarchical organization of gamma and theta oscillatory dynamics in schizophrenia. Biol Psychiatry 2012; 71(10): 873–880, https://doi.org/10.1016/j.biopsych.2012.01.016.
- Froriep U.P., Kumar A., Cosandier-Rimélé D., Häussler U., Kilias A., Haas C.A., Egert U. Altered theta coupling between medial entorhinal cortex and dentate gyrus in temporal lobe epilepsy. Epilepsia 2012; 53(11): 1937–1947, https://doi.org/10.1111/j.1528-1167.2012.03662.x.
- Bakker A., Krauss G.L., Albert M.S., Speck C.L., Jones L.R., Stark C.E., Yassa M.A., Bassett S.S., Shelton A.L., Gallagher M. Reduction of hippocampal hyperactivity improves cognition in amnestic mild cognitive impairment. Neuron 2012; 74(3): 467–474, https://doi.org/10.1016/j.neuron.2012.03.023.
- Inostroza M., Brotons-Mas J.R., Laurent F., Cid E., de la Prida L.M. Specific impairment of “what-where-when” episodic-like memory in experimental models of temporal lobe epilepsy. J Neurosci 2013; 33(45): 17749–17762, https://doi.org/10.1523/jneurosci.0957-13.2013.
- Laurent F., Brotons-Mas J.R., Cid E., Lopez-Pigozzi D., Valero M., Gal B., de la Prida L.M. Proximodistal structure of theta coordination in the dorsal hippocampus of epileptic rats. J Neurosci 2015; 35(11): 4760–4775, https://doi.org/10.1523/jneurosci.4297-14.2015.
- Kitchigina V.F., Butuzova M.V. Theta activity of septal neurons during different epileptic phases: the same frequency but different significance? Exp Neurol 2009; 216(2): 449–458, https://doi.org/10.1016/j.expneurol.2009.01.001.
- Rodriguez E., George N., Lachaux J.P., Martinerie J., Renault B., Varela F.J. Perception’s shadow: long-distance synchronization of human brain activity. Nature 1999; 397(6718): 430–433, https://doi.org/10.1038/17120.
- Hurtado J.M., Rubchinsky L.L., Sigvardt K.A. Statistical method for detection of phase-locking episodes in neural oscillations. J Neurophysiol 2004; 91(4): 1883–1898, https://doi.org/10.1152/jn.00853.2003.
- Canolty R.T., Edwards E., Dalal S.S., Soltani M., Nagarajan S.S., Kirsch H.E., Berger M.S., Barbaro N.M., Knight R.T. High gamma power is phase-locked to theta oscillations in human neocortex. Science 2006; 313(5793): 1626–1628, https://doi.org/10.1126/science.1128115.
- Axmacher N., Henseler M.M., Jensen O., Weinreich I., Elger C.E., Fell J. Cross-frequency coupling supports multi-item working memory in the human hippocampus. Proc Natl Acad Sci U S A 2010; 107(7): 3228–3233, https://doi.org/10.1073/pnas.0911531107.
- Montgomery S.M., Buzsaki G. Gamma oscillations dynamically couple hippocampal CA3 and CA1 regions during memory task performance. Proc Natl Acad Sci U S A 2007; 104(36): 14495–14500, https://doi.org/10.1073/pnas.0701826104.
- Tort A.B., Kramer M.A., Thorn C., Gibson D.J., Kubota Y., Graybiel A.M., Kopell N.J. Dynamic cross-frequency couplings of local field potential oscillations in rat striatum and hippocampus during performance of a T-maze task. Proc Natl Acad Sci U S A 2008; 105(51): 20517–20522, https://doi.org/10.1073/pnas.0810524105.
- Nacher V., Ledberg A., Deco G., Romo R. Coherent delta-band oscillations between cortical areas correlate with decision making. Proc Natl Acad Sci U S A 2013; 110(37): 15085–15090, https://doi.org/10.1073/pnas.1314681110.
- Wulff P., Ponomarenko A.A., Bartos M., Korotkova T.M., Fuchs E.C., Bähner F., Both M., Tort A.B., Kopell N.J., Wisden W., Monyer H. Hippocampal theta rhythm and its coupling with gamma oscillations require fast inhibition onto parvalbumin-positive interneurons. Proc Natl Acad Sci U S A 2009; 106(9): 3561–3566, https://doi.org/10.1073/pnas.0813176106.
- Siegel M., Warden M.R., Miller E.K. Phase-dependent neuronal coding of objects in short-term memory. Proc Natl Acad Sci U S A 2009; 106(50): 21341–21346, https://doi.org/10.1073/pnas.0908193106.
- Gregoriou G.G., Gotts S.J., Zhou H., Desimone R. High-frequency, long-range coupling between prefrontal and visual cortex during attention. Science 2009; 324(5931): 1207–1210, https://doi.org/10.1126/science.1171402.
- Palva J.M. Phase synchrony among neuronal oscillations in the human cortex. J Neurosci 2005; 25(15): 3962–3972, https://doi.org/10.1523/jneurosci.4250-04.2005.
- Palva J.M., Monto S., Kulashekhar S., Palva S. Neuronal synchrony reveals working memory networks and predicts individual memory capacity. Proc Natl Acad Sci U S A 2010; 107(16): 7580–7585, https://doi.org/10.1073/pnas.0913113107.
- Holz E.M., Glennon M., Prendergast K., Sauseng P. Theta–gamma phase synchronization during memory matching in visual working memory. NeuroImage 2010; 52(1): 326–335, https://doi.org/10.1016/j.neuroimage.2010.04.003.
- Shirvalkar P.R., Rapp P.R., Shapiro M.L. Bidirectional changes to hippocampal theta-gamma comodulation predict memory for recent spatial episodes. Proc Natl Acad Sci U S A 2010; 107(15): 7054–7059, https://doi.org/10.1073/pnas.0911184107.
- Tanninen S.E., Nouriziabari B., Morrissey M.D., Bakir R., Dayton R.D., Klein R.L., Takehara-Nishiuchi K. Entorhinal tau pathology disrupts hippocampal-prefrontal oscillatory coupling during associative learning. Neurobiol Aging 2017; 58: 151–162, https://doi.org/10.1016/j.neurobiolaging.2017.06.024.
- Friston K.J. Another neural code? NeuroImage 1997; 5(3): 213–220, https://doi.org/10.1006/nimg.1997.0260.
- Buzsáki G., Lai-Wo S.L., Vanderwolf C.H. Cellular bases of hippocampal EEG in the behaving rat. Brain Res 1983; 6(2): 139–171, https://doi.org/10.1016/0165-0173(83)90037-1.
- Buzsáki G., Buhl D.L., Harris K.D., Csicsvari J., Czéh B., Morozov A. Hippocampal network patterns of activity in the mouse. Neuroscience 2003; 116(1): 201–211, https://doi.org/10.1016/s0306-4522(02)00669-3.
- Soltesz I., Deschenes M. Low- and high-frequency membrane potential oscillations during theta activity in CA1 and CA3 pyramidal neurons of the rat hippocampus under ketamine-xylazine anesthesia. J Neurophysiol 1993; 70(1): 97–116, https://doi.org/10.1152/jn.1993.70.1.97.
- Lisman J., Idiart M. Storage of 7 +/– 2 short-term memories in oscillatory subcycles. Science 1995; 267(5203): 1512–1515, https://doi.org/10.1126/science.7878473.
- Mormann F., Fell J., Axmacher N., Weber B., Lehnertz K., Elger C.E., Fernández G. Phase/amplitude reset and theta-gamma interaction in the human medial temporal lobe during a continuous word recognition memory task. Hippocampus 2005; 15(7): 890–900, https://doi.org/10.1002/hipo.20117.
- Sirota A., Montgomery S., Fujisawa S., Isomura Y., Zugaro M., Buzsáki G. Entrainment of neocortical neurons and gamma oscillations by the hippocampal theta rhythm. Neuron 2008; 60(4): 683–697, https://doi.org/10.1016/j.neuron.2008.09.014.
- Tort A.B.L., Komorowski R., Eichenbaum H., Kopell N. Measuring phase-amplitude coupling between neuronal oscillations of different frequencies. J Neurophysiol 2010; 104(2): 1195–1210, https://doi.org/10.1152/jn.00106.2010.
- Scheffer-Teixeira R., Belchior H., Caixeta F.V., Souza B.C., Ribeiro S., Tort A.B.L. Theta phase modulates multiple layer-specific oscillations in the CA1 region. Cereb Cortex 2011; 22(10): 2404–2414, https://doi.org/10.1093/cercor/bhr319.
- Schomburg E.W., Fernández-Ruiz A., Mizuseki K., Berényi A., Anastassiou C.A., Koch C., Buzsáki G. Theta phase segregation of input-specific gamma patterns in entorhinal-hippocampal networks. Neuron 2014; 84(2): 470–485, https://doi.org/10.1016/j.neuron.2014.08.051.
- Tass P., Rosenblum M.G., Weule J., Kurths J., Pikovsky A., Volkmann J., Schnitzler A., Freund H.-J. Detection of n:m phase locking from noisy data: application to magnetoencephalography. Phys Rev Lett 1998; 81(15): 3291–3294.
- Belluscio M.A., Mizuseki K., Schmidt R., Kempter R., Buzsaki G. Cross-frequency phase-phase coupling between theta and gamma oscillations in the hippocampus. J Neurosci 2012; 32(2): 423–435, https://doi.org/10.1523/jneurosci.4122-11.2012.
- Zheng C., Zhang T. Alteration of phase–phase coupling between theta and gamma rhythms in a depression-model of rats. Cogn Neurodyn 2012; 7(2): 167–172, https://doi.org/10.1007/s11571-012-9225-x.
- Xu X., Liu C., Li Z., Zhang T. Effects of hydrogen sulfide on modulation of theta–gamma coupling in hippocampus in vascular dementia rats. Brain Topogr 2015; 28(6): 879–894, https://doi.org/10.1007/s10548-015-0430-x.
- Zheng C., Zhang T. Synaptic plasticity-related neural oscillations on hippocampus–prefrontal cortex pathway in depression. Neuroscience 2015; 292: 170–180, https://doi.org/10.1016/j.neuroscience.2015.01.071.
- Ittner A.A., Gladbach A., Bertz J., Suh L.S., Ittner L.M. p38 MAP kinase-mediated NMDA receptor-dependent suppression of hippocampal hypersynchronicity in a mouse model of Alzheimer’s disease. Acta Neuropathol Commun 2014; 2(1): 149, https://doi.org/10.1186/s40478-014-0149-z.
- Quilichini P., Sirota A., Buzsaki G. Intrinsic circuit organization and theta-gamma oscillation dynamics in the entorhinal cortex of the rat. J Neurosci 2010; 30(33): 11128–11142, https://doi.org/10.1523/jneurosci.1327-10.2010.
- Szczepanski S.M., Crone N.E., Kuperman R.A., Auguste K.I., Parvizi J., Knight R.T. Dynamic changes in phase-amplitude coupling facilitate spatial attention control in fronto-parietal cortex. PLoS Biology 2014; 12(8): e1001936, https://doi.org/10.1371/journal.pbio.1001936.
- Buzsáki G. Rhythms of the brain. Oxford University Press; 2006, https://doi.org/10.1093/acprof:oso/9780195301069.001.0001.
- Buzsáki G., Mizuseki K. The log-dynamic brain: how skewed distributions affect network operations. Nat Rev Neurosci 2014; 15(4): 264–278, https://doi.org/10.1038/nrn3687.
- Knight R.T. Neuroscience: neural networks debunk phrenology. Science 2007; 316(5831): 1578–1579, https://doi.org/10.1126/science.1144677.
- Haider B., McCormick D.A. Rapid neocortical dynamics: cellular and network mechanisms. Neuron 2009; 62(2): 171–189, https://doi.org/10.1016/j.neuron.2009.04.008.
- Voytek B., D’Esposito M., Crone N., Knight R.T. A method for event-related phase/amplitude coupling. NeuroImage 2013; 64: 416–424, https://doi.org/10.1016/j.neuroimage.2012.09.023.
- Lega B.C., Jacobs J., Kahana M. Human hippocampal theta oscillations and the formation of episodic memories. Hippocampus 2011; 22(4): 748–761, https://doi.org/10.1002/hipo.20937.
- Newman E.L., Gillet S.N., Climer J.R., Hasselmo M.E. Cholinergic blockade reduces theta–gamma phase amplitude coupling and speed modulation of theta frequency consistent with behavioral effects on encoding. J Neurosci 2013; 33(50): 19635–19646, https://doi.org/10.1523/jneurosci.2586-13.2013.
- Schack B., Weiss S. Quantification of phase synchronization phenomena and their importance for verbal memory processes. Biol Cybern 2005; 92(4): 275–287, https://doi.org/10.1007/s00422-005-0555-1.
- Fell J., Axmacher N. The role of phase synchronization in memory processes. Nat Rev Neurosci 2011; 12(2): 105–118, https://doi.org/10.1038/nrn2979.
- Scheffer-Teixeira R., Tort A.B. On cross-frequency phase-phase coupling between theta and gamma oscillations in the hippocampus. Elife 2016; 5: e20515, https://doi.org/10.7554/elife.20515.
- Markram H. Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science 1997; 275(5297): 213–215, https://doi.org/10.1126/science.275.5297.213.
- Morimoto K., Fahnestock M., Racine R.J. Kindling and status epilepticus models of epilepsy: rewiring the brain. Prog Neurobiol 2004; 73(1): 1–60, https://doi.org/10.1016/j.pneurobio.2004.03.009.
- Dupont S., Van de Moortele P.F., Samson S., Hasboun D., Poline J.B., Adam C., Lehéricy S., Le Bihan D., Samson Y., Baulac M. Episodic memory in left temporal lobe epilepsy: a functional MRI study. Brain 2000; 123(Pt 8): 1722–1732, https://doi.org/10.1093/brain/123.8.1722.
- Helmstaedter C. Effects of chronic epilepsy on declarative memory systems. Prog Brain Res 2002; 135: 439–453, https://doi.org/10.1016/s0079-6123(02)35041-6.
- Burgess N., Maguire E.A., O’Keefe J. The human hippocampus and spatial and episodic memory. Neuron 2002; 35(4): 625–641, https://doi.org/10.1016/s0896-6273(02)00830-9.
- Tulving E. Episodic memory: from mind to brain. Annu Rev Psychol 2002; 53(1): 1–25, https://doi.org/10.1146/annurev.psych.53.100901.135114.
- Lega B., Dionisio S., Bingaman W., Najm I., Gonzalez-Martinez J. The gamma band effect for episodic memory encoding is absent in epileptogenic hippocampi. Clin Neurophysiol 2015; 126(5): 866–872, https://doi.org/10.1016/j.clinph.2014.07.035.
- O’Keefe J., Nadel L. The hippocampus as a cognitive map. Oxford, UK: Oxford University Press; 1978.
- Komorowski R.W., Manns J.R., Eichenbaum H. Robust conjunctive item-place coding by hippocampal neurons parallels learning what happens where. J Neurosci 2009; 29(31): 9918–9929, https://doi.org/10.1523/jneurosci.1378-09.2009.
- Mankin E.A., Sparks F.T., Slayyeh B., Sutherland R.J., Leutgeb S., Leutgeb J.K. Neuronal code for extended time in the hippocampus. Proc Natl Acad Sci U S A 2012; 109(47): 19462–19467, https://doi.org/10.1073/pnas.1214107109.
- Kraus B.J., Robinson R.J., White J.A., Eichenbaum H., Hasselmo M.E. Hippocampal “time cells”: time versus path integration. Neuron 2013; 78(6): 1090–1101, https://doi.org/10.1016/j.neuron.2013.04.015.
- Kitamura T., Pignatelli M., Suh J., Kohara K., Yoshiki A., Abe K., Tonegawa S. Island cells control temporal association memory. Science 2014; 343(6173): 896–901, https://doi.org/10.1126/science.1244634.
- Frank L.M., Brown E.N., Wilson M. Trajectory encoding in the hippocampus and entorhinal cortex. Neuron 2000; 27(1): 169–178, https://doi.org/10.1016/s0896-6273(00)00018-0.
- Wood E.R., Dudchenko P.A., Robitsek R.J., Eichenbaum H. Hippocampal neurons encode information about different types of memory episodes occurring in the same location. Neuron 2000; 27(3): 623–633, https://doi.org/10.1016/s0896-6273(00)00071-4.
- Huxter J.R., Senior T.J., Allen K., Csicsvari J. Theta phase–specific codes for two-dimensional position, trajectory and heading in the hippocampus. Nat Neurosci 2008; 11(5): 587–594, https://doi.org/10.1038/nn.2106.
- Mizuseki K., Sirota A., Pastalkova E., Buzsáki G. Theta oscillations provide temporal windows for local circuit computation in the entorhinal-hippocampal loop. Neuron 2009; 64(2): 267–280, https://doi.org/10.1016/j.neuron.2009.08.037.
- Easton A., Douchamps V., Eacott M., Lever C. A specific role for septohippocampal acetylcholine in memory? Neuropsychologia 2012; 50(13): 3156–3168, https://doi.org/10.1016/j.neuropsychologia.2012.07.022.
- Buzsáki G., Moser E.I. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat Neurosci 2013; 16(2): 130–138, https://doi.org/10.1038/nn.3304.
- Siapas A.G., Lubenov E.V., Wilson M.A. Prefrontal phase locking to hippocampal theta oscillations. Neuron 2005; 46(1): 141–151, https://doi.org/10.1016/j.neuron.2005.02.028.
- Jones M.W., Wilson M.A. Theta rhythms coordinate hippocampal–prefrontal interactions in a spatial memory task. PLoS Biology 2005; 3(12): e402, https://doi.org/10.1371/journal.pbio.0030402.
- Broggini A.C.S., Esteves I.M., Romcy-Pereira R.N., Leite J.P., Leão R.N. Pre-ictal increase in theta synchrony between the hippocampus and prefrontal cortex in a rat model of temporal lobe epilepsy. Exp Neurol 2016; 279: 232–242, https://doi.org/10.1016/j.expneurol.2016.03.007.
- Arabadzisz D., Antal K., Parpan F., Emri Z., Fritschy J.-M. Epileptogenesis and chronic seizures in a mouse model of temporal lobe epilepsy are associated with distinct EEG patterns and selective neurochemical alterations in the contralateral hippocampus. Exp Neurol 2005; 194(1): 76–90, https://doi.org/10.1016/j.expneurol.2005.01.029.
- Colom L.V., García-Hernández A., Castañeda M.T., Perez-Cordova M.G., Garrido-Sanabria E.R. Septo-hippocampal networks in chronically epileptic rats: potential antiepileptic effects of theta rhythm generation. J Neurophysiol 2006; 95(6): 3645–3653, https://doi.org/10.1152/jn.00040.2006.
- Dugladze T., Vida I., Tort A.B., Gross A., Otahal J., Heinemann U., Kopell N.J., Gloveli T. Impaired hippocampal rhythmogenesis in a mouse model of mesial temporal lobe epilepsy. Proc Natl Acad Sci U S A 2007; 104(44): 17530–17535, https://doi.org/10.1073/pnas.0708301104.
- Marcelin B., Chauvière L., Becker A., Migliore M., Esclapez M., Bernard C. h channel-dependent deficit of theta oscillation resonance and phase shift in temporal lobe epilepsy. Neurobiol Dis 2009; 33(3): 436–447, https://doi.org/10.1016/j.nbd.2008.11.019.
- García-Hernández A., Bland B.H., Facelli J.C., Colom L.V. Septo-hippocampal networks in chronic epilepsy. Exp Neurol 2010; 222(1): 86–92, https://doi.org/10.1016/j.expneurol.2009.12.010.
- Selkoe D.J. Toward a comprehensive theory for Alzheimer’s disease. Hypothesis: Alzheimer’s disease is caused by the cerebral accumulation and cytotoxicity of amyloid β-protein. Ann N Y Acad Sci 2006; 924(1): 17–25, https://doi.org/10.1111/j.1749-6632.2000.tb05554.x.
- Palop J.J., Mucke L. Amyloid-beta-induced neuronal dysfunction in Alzheimer’s disease: from synapses toward neural networks. Nat Neurosci 2010; 13(7): 812–818, https://doi.org/10.1038/nn.2583.
- Francis B.M., Kim J., Barakat M.E., Fraenkl S., Yücel Y.H., Peng S., Michalski B., Fahnestock M., McLaurin J., Mount H.T. Object recognition memory and BDNF expression are reduced in young TgCRND8 mice. Neurobiol Aging 2012; 33(3): 555–563, https://doi.org/10.1016/j.neurobiolaging.2010.04.003.
- Kametani F., Hasegawa M. Reconsideration of amyloid hypothesis and tau hypothesis in Alzheimer’s disease. Front Neurosci 2018; 12: 25–36, https://doi.org/10.3389/fnins.2018.00025.
- Gureviciene I., Gurevicius K., Mugantseva E., Kislin M., Khiroug L., Tanila H. Amyloid plaques show binding capacity of exogenous injected amyloid-β. J Alzheimers Dis 2016; 55(1): 147–157, https://doi.org/10.3233/jad-160453.
- Didic M., Barbeau E.J., Felician O., Tramoni E., Guedj E., Poncet M., Ceccaldi M. Which memory system is impaired first in Alzheimer’s disease? J Alzheimers Dis 2011; 27(1): 11–22, https://doi.org/10.3233/jad-2011-110557.
- Lithfous S., Dufour A., Després O. Spatial navigation in normal aging and the prodromal stage of Alzheimer’s disease: insights from imaging and behavioral studies. Ageing Research Reviews 2013; 12(1): 201–213, https://doi.org/10.1016/j.arr.2012.04.007.
- Adler G., Brassen S., Jajcevic A. EEG coherence in Alzheimer’s dementia. J Neural Transm 2003; 110(9): 1051–1058, https://doi.org/10.1007/s00702-003-0024-8.
- Herrmann C., Demiralp T. Human EEG gamma oscillations in neuropsychiatric disorders. Clin Neurophysiol 2005; 116(12): 2719–2733, https://doi.org/10.1016/j.clinph.2005.07.007.
- van der Hiele K., Vein A.A., van der Welle A., van der Grond J., Westendorp R.G., Bollen E.L., van Buchem M.A., van Dijk J.G., Middelkoop H.A. EEG and MRI correlates of mild cognitive impairment and Alzheimer’s disease. Neurobiol Aging 2007; 28(9): 1322–1329, https://doi.org/10.1016/j.neurobiolaging.2006.06.006.
- Czigler B., Csikós D., Hidasi Z., Anna Gaál Z., Csibri E., Kiss E., Salacz P., Molnár M. Quantitative EEG in early Alzheimer’s disease patients — power spectrum and complexity features. Int J Psychophysiol 2008; 68(1): 75–80, https://doi.org/10.1016/j.ijpsycho.2007.11.002.
- Moretti D.V., Pievani M., Geroldi C., Binetti G., Zanetti O., Rossini P.M., Frisoni G.B. EEG markers discriminate among different subgroup of patients with mild cognitive impairment. Am J Alzheimers Dis Other Demen 2010; 25(1): 58–73, https://doi.org/10.1177/1533317508329814.
- Caravaglios G., Castro G., Costanzo E., Di Maria G., Mancuso D., Muscoso E.G. Theta power responses in mild Alzheimer’s disease during an auditory oddball paradigm: lack of theta enhancement during stimulus processing. J Neural Transm 2010; 117(10): 1195–1208, https://doi.org/10.1007/s00702-010-0488-2.
- Wang J., Fang Y., Wang X., Yang H., Yu X., Wang H. Enhanced gamma activity and cross-frequency interaction of resting-state electroencephalographic oscillations in patients with Alzheimer’s disease. Front Aging Neurosci 2017; 9: 243, https://doi.org/10.3389/fnagi.2017.00243.
- Schneider F., Baldauf K., Wetzel W., Reymann K.G. Behavioral and EEG changes in male 5xFAD mice. Physiol Behav 2014; 135: 25–33, https://doi.org/10.1016/j.physbeh.2014.05.041.
- Goodman M.S., Kumar S., Zomorrodi R., Ghazala Z., Cheam A.S.M., Barr M.S., Daskalakis Z.J., Blumberger D.M., Fischer C., Flint A., Mah L., Herrmann N., Bowie C.R., Mulsant B.H., Rajji T.K. Theta-gamma coupling and working memory in Alzheimer’s dementia and mild cognitive impairment. Front Aging Neurosci 2018; 10: 101, https://doi.org/10.3389/fnagi.2018.00101.
- Goutagny R., Gu N., Cavanagh C., Jackson J., Chabot J.G., Quirion R., Krantic S., Williams S. Alterations in hippocampal network oscillations and theta-gamma coupling arise before Aβ overproduction in a mouse model of Alzheimer’s disease. Eur J Neurosci 2013; 37(12): 1896–1902, https://doi.org/10.1111/ejn.12233.
- Scott L., Feng J., Kiss T., Needle E., Atchison K., Kawabe T.T., Milici A.J., Hajós-Korcsok E., Riddell D., Hajós M. Age-dependent disruption in hippocampal theta oscillation in amyloid-β overproducing transgenic mice. Neurobiol Aging 2012; 33(7): 1481.e13–1481.e23, https://doi.org/10.1016/j.neurobiolaging.2011.12.010.
- Seabrook G.R., Smith D.W., Bowery B.J., Easter A., Reynolds T., Fitzjohn S.M., Morton R.A., Zheng H., Dawson G.R., Sirinathsinghji D.J., Davies C.H., Collingridge G.L., Hill R.G. Mechanisms contributing to the deficits in hippocampal synaptic plasticity in mice lacking amyloid precursor protein. Neuropharmacology 1999; 38(3): 349–359, https://doi.org/10.1016/s0028-3908(98)00204-4.
- Senechal Y., Kelly P.H., Dev K.K. Amyloid precursor protein knockout mice show age-dependent deficits in passive avoidance learning. Behav Brain Res 2008; 186(1): 126–132, https://doi.org/10.1016/j.bbr.2007.08.003.
- Zhang X., Zhong W., Brankačk J., Weyer S.W., Müller U.C., Tort A.B., Draguhn A. Impaired theta-gamma coupling in APP-deficient mice. Sci Rep 2016; 6: 21948, https://doi.org/10.1038/srep21948.
- Armon C., Peterson G.W., Liwnicz B.H. Alzheimer’s disease underlies some cases of complex partial status epilepticus. J Clin Neurophysiol 2000; 17(5): 511–518, https://doi.org/10.1097/00004691-200009000-00011.
- Amatniek J.C., Hauser W.A., DelCastillo-Castaneda C., Jacobs D.M., Marder K., Bell K., Albert M., Brandt J., Stern Y. Incidence and predictors of seizures in patients with Alzheimer’s disease. Epilepsia 2006; 47(5): 867–872, https://doi.org/10.1111/j.1528-1167.2006.00554.x.
- Braak H., Braak E. Evolution of the neuropathology of Alzheimer’s disease. Acta Neurol Scand Suppl 1996; 165: 3–12, https://doi.org/10.1111/j.1600-0404.1996.tb05866.x.
- Born H.A., Kim J.Y., Savjani R.R., Das P., Dabaghian Y.A., Guo Q., Yoo J.W., Schuler D.R., Cirrito J.R., Zheng H., Golde T.E., Noebels J.L., Jankowsky J.L. Genetic suppression of transgenic APP rescues hypersynchronous network activity in a mouse model of Alzeimer’s disease. J Neurosci 2014; 34(11): 3826–3840, https://doi.org/10.1523/jneurosci.5171-13.2014.
- Chan J., Jones N.C., Bush A.I., O’Brien T.J., Kwan P. A mouse model of Alzheimer’s disease displays increased susceptibility to kindling and seizure-associated death. Epilepsia 2015; 56(6): e73–e77, https://doi.org/10.1111/epi.12993.
- Bird S.B., Orr P.G., Mazzola J.L., Brush D.E., Boyer E.W. Levofloxacin-related seizure activity in a patient with Alzheimer’s disease: assessment of potential risk factors. J Clin Psychopharmacol 2005; 25(3): 287–288, https://doi.org/10.1097/01.jcp.0000162811.15066.8e.
- Bernardi S., Scaldaferri N., Vanacore N., Trebbastoni A., Francia A., D’Amico A., Prencipe M. Seizures in Alzheimer’s disease: a retrospective study of a cohort of outpatients. Epileptic Disord 2010; 12(1): 16–21.
- Palop J.J., Mucke L. Network abnormalities and interneuron dysfunction in Alzheimer disease. Nat Rev Neurosci 2016; 17(12): 777–792, https://doi.org/10.1038/nrn.2016.141.
- Bezzina C., Verret L., Juan C., Remaud J., Halley H., Rampon C., Dahan L. Early onset of hypersynchronous network activity and expression of a marker of chronic seizures in the Tg2576 mouse model of Alzheimer’s disease. PLoS One 2015; 10(3): e0119910, https://doi.org/10.1371/journal.pone.0119910.
- Palop J.J., Chin J., Roberson E.D., Wang J., Thwin M.T., Bien-Ly N., Yoo J., Ho K.O., Yu G.Q., Kreitzer A., Finkbeiner S., Noebels J.L., Mucke L. Aberrant excitatory neuronal activity and compensatory remodeling of inhibitory hippocampal circuits in mouse models of Neuron 2007; 55(5): 697–711, https://doi.org/10.1016/j.neuron.2007.07.025.
- Bero A.W., Yan P., Roh J.H., Cirrito J.R., Stewart F.R., Raichle M.E., Lee J.M., Holtzman D.M. Neuronal activity regulates the regional vulnerability to amyloid-β deposition. Nat Neurosci 2011; 14(6): 750–756, https://doi.org/10.1038/nn.2801.
- Cirrito J.R., Kang J.-E., Lee J., Stewart F.R., Verges D.K., Silverio L.M., Bu G., Mennerick S., Holtzman D.M. Endocytosis is required for synaptic activity-dependent release of amyloid-β in vivo. Neuron 2008; 58(1): 42–51, https://doi.org/10.1016/j.neuron.2008.02.003.
- Gallassi R. Epileptic amnesic syndrome: an update and further considerations. Epilepsia 2006; 47(s2): 103–105, https://doi.org/10.1111/j.1528-1167.2006.00704.x.
- Vossel K.A., Beagle A.J., Rabinovici G.D., Shu H., Lee S.E., Naasan G., Hegde M., Cornes S.B., Henry M.L., Nelson A.B., Seeley W.W., Geschwind M.D., Gorno-Tempini M.L., Shih T., Kirsch H.E., Garcia P.A., Miller B.L., Mucke L. Seizures and epileptiform activity in the early stages of Alzheimer disease. JAMA Neurol 2013; 70(9): 1158, https://doi.org/10.1001/jamaneurol.2013.136.
- Cretin B., Sellal F., Philippi N., Bousiges O., Di Bitonto L., Martin-Hunyadi C., Blanc F. Epileptic prodromal Alzheimer’s disease, a retrospective study of 13 new cases: expanding the spectrum of Alzheimer’s disease to an epileptic variant? J Alzheimers Dis 2016; 52(3): 1125–1133, https://doi.org/10.3233/jad-150096.
- Chin J., Scharfman H.E. Shared cognitive and behavioral impairments in epilepsy and Alzheimer’s disease and potential underlying mechanisms. Epilepsy Behav 2013; 26(3): 343–351, https://doi.org/10.1016/j.yebeh.2012.11.040.
- Eichler S.A. E-I balance and human diseases — from molecules to networking. Front Mol Neurosci 2008; 1: 2, https://doi.org/10.3389/neuro.02.002.2008.
- Noebels J. A perfect storm: converging paths of epilepsy and Alzheimer’s dementia intersect in the hippocampal formation. Epilepsia 2011; 52: 39–46, https://doi.org/10.1111/j.1528-1167.2010.02909.x.
- Saito Y., Inoue T., Zhu G., Kimura N., Okada M., Nishimura M., Kimura N., Murayama S., Kaneko S., Shigemoto R., Imoto K., Suzuki T. Hyperpolarization-activated cyclic nucleotide gated channels: a potential molecular link between epileptic seizures and Aβ generation in Alzheimer’s disease. Mol Neurodegener 2012; 7: 50, https://doi.org/10.1186/1750-1326-7-50.
- Varga E., Juhász G., Bozsó Z., Penke B., Fülöp L., Szegedi V. Abeta(1-42) enhances neuronal excitability in the CA1 via NR2B subunit-containing NMDA receptors. Neural Plast 2014; 2014: 1–12, https://doi.org/10.1155/2014/584314.
- Irizarry M.C., Jin S., He F., Emond J.A., Raman R., Thomas R.G., Sano M., Quinn J.F., Tariot P.N., Galasko D.R., Ishihara L.S., Weil J.G., Aisen P.S. Incidence of new-onset seizures in mild to moderate Alzheimer disease. Arch Neurol 2012; 69(3): 368, https://doi.org/10.1001/archneurol.2011.830.
- Scarmeas N., Honig L.S., Choi H., Cantero J., Brandt J., Blacker D., Albert M., Amatniek J.C., Marder K., Bell K., Hauser W.A., Stern Y. Seizures in Alzheimer disease: who, when, and how common? Arch Neurol 2009; 66(8): 992–997, https://doi.org/10.1001/archneurol.2009.130.
- Sarkis R.A., Willment K.C., Gale S.A., Dworetzky B.A. Recurrent epileptic auras as a presenting symptom of Alzheimer’s disease. Front Neurol 2017; 8: 360, https://doi.org/10.3389/fneur.2017.00360.
- Minkeviciene R., Rheims S., Dobszay M.B., Zilberter M., Hartikainen J., Fülöp L., Penke B., Zilberter Y., Harkany T., Pitkänen A., Tanila H. Amyloid beta-induced neuronal hyperexcitability triggers progressive epilepsy. J Neurosci 2009; 29(11): 3453–3462, https://doi.org/10.1523/jneurosci.5215-08.2009.
- Vogt D.L., Thomas D., Galvan V., Bredesen D.E., Lamb B.T., Pimplikar S.W. Abnormal neuronal networks and seizure susceptibility in mice overexpressing the APP intracellular domain. Neurobiol Aging 2011; 32(9): 1725–1729, https://doi.org/10.1016/j.neurobiolaging.2009.09.002.
- Yener G.G., Başar E. Biomarkers in Alzheimer’s disease with a special emphasis on event-related oscillatory responses. Suppl Clin Neurophysiol 2013: 62: 237–273, https://doi.org/10.1016/b978-0-7020-5307-8.00020-x.
- Wang B., Meng L. Functional brain network alterations in epilepsy: a magnetoencephalography study. Epilepsy Res 2016; 126: 62–69, https://doi.org/10.1016/j.eplepsyres.2016.06.014.








