Активность зеркальных нейронов у человека при наблюдении и восприятии времени
Цель исследования — изучение активности зеркальных нейронов у человека при наблюдении и восприятии им коротких интервалов времени в зависимости от латеральной организации мозга.
Материалы и методы. В исследованиях участвовали добровольцы — юноши в возрасте 18–27 лет. В ходе предварительного обследования выявляли особенности латеральной организации мозга с определением ведущей руки (методом анкетирования) и речевого полушария (с помощью дихотического теста). При наблюдении и отмеривании коротких интервалов времени, а также при наблюдении и репродукции пятисекундного ритма регистрировали ЭЭГ в лобных, центральных, височных, теменных и затылочных отведениях по системе 10–20%. С целью изучения пространственной локализации зеркальных нейронов в части опытов при наблюдении и восприятии времени исследовали активность мозга с помощью функциональной МРТ (фМРТ). В качестве ЭЭГ-маркеров активации зеркальных нейронов использовали депрессию мю-ритма и корковые взаимодействия на частоте этого ритма между центральными и другими зонами коры.
Результаты. Установлено, что наблюдение и выполнение деятельности, связанной с восприятием времени испытуемыми, сопровождаются депрессией мю-ритма ЭЭГ и чаще всего усилением уровней корковых связей на частоте этого ритма. Характер этих изменений зависит от частоты мю-ритма, латеральной организации мозга, от вида и этапа выполняемой деятельности. Результаты фМРТ-сканирования мозга при наблюдении и восприятии времени показали, что в процессах восприятия времени фактически участвуют две системы, работающие совместно. Это система зеркальных нейронов, которая включает зоны премоторной, моторной, сенсомоторной и дополнительной моторной коры, а также области скорлупы, хвостатого ядра, супрамаргинальной извилины, височные отделы мозга; и система отсчета времени, включающая области поясной извилины, левого таламуса, височные отделы мозга, зрительную кору, области предклинья и области мозжечка.
Результаты исследования имеют важное теоретическое значение для понимания роли зеркальных нейронов в процессах восприятия времени.
Введение
Изучение роли зеркальных нейронов в когнитивных процессах имеет важное значение для понимания социального поведения человека. Согласно популярной в настоящее время гипотезе [1], зеркальные нейроны могут служить нейрональной основой для интерпретации действий, подражательного обучения и имитации поведения других людей. По мнению некоторых исследователей, нарушение функций зеркальных нейронов может быть причиной аутизма [2].
Считается, что характерным ЭЭГ-коррелятом активации зеркальных нейронов является депрессия мю-ритма частотой 8–13 Гц, который регистрируется в центральных областях коры и не подавляется при зрительной и слуховой стимуляции [3]. Вместе с тем проведенные в этом направлении исследования дали неоднозначные результаты. В частности, при изучении электрической активности мозга в центральных и затылочных отведениях в процессе наблюдения видеозаписей движений рук и калейдоскопического паттерна на экране монитора авторы приходят к выводу, что подавление мю-ритма может быть использовано как индикатор активации зеркальных нейронов человека, но эффект — слабый и ненадежный и легко смешивается с подавлением затылочного альфа-ритма [4]. Кроме того, некоторые исследователи [5] выделяют в мю-ритме два поддиапазона: 8–10 и 10–12 Гц, которые, по их мнению, функционально различаются. В ряде работ обнаружены значительные индивидуальные различия реактивности мю-ритма при выполнении двигательных задач [6–9].
Все эти данные указывают на необходимость дальнейших исследований роли зеркальных нейронов в когнитивных процессах и, в частности, в процессах восприятия времени, в которых их функция практически не изучалась. По-прежнему актуальны исследования, направленные на поиск надежных ЭЭГ-маркеров активации этих нейронов, на изучение зависимости их активности от индивидуальных особенностей человека, на уточнение их пространственной локализации.
Цель исследования — изучение активности зеркальных нейронов у человека при наблюдении и восприятии им коротких интервалов времени в зависимости от латеральной организации мозга.
Материалы и методы
Для достижения поставленной цели проведены ЭЭГ- и фМРТ-исследования активности мозга у юношей при наблюдении и восприятии ими коротких интервалов времени.
В ЭЭГ-исследовании участвовали добровольцы, практически здоровые юноши — студенты в возрасте от 18 до 23 лет (n=31). Все испытуемые дали информированное согласие на участие. Исследование проведено в соответствии с Хельсинкской декларацией (2013) и разрешено комиссией по биоэтике Биологического института Томского государственного университета.
В ходе предварительного обследования с помощью стандартных методов изучали особенности латеральной организации мозга с определением ведущей руки (методом анкетирования) и речевого полушария (с помощью дихотического теста). По результатам анкетирования подсчитывали в баллах показатель мануального предпочтения и коэффициент правого уха.
Для решения поставленных задач проведено 6 серий наблюдений. В первой серии («наблюдение за отмериванием длительности») испытуемый наблюдал за оператором, который средним и указательным пальцами ведущей руки отмеривал короткие интервалы времени паузой между двумя нажатиями на клавишу «пробел». Длительность интервалов времени задавалась зрительными стимулами (светлый квадрат, появляющийся через 200 или 800 мс в центре затемненного экрана монитора). Стимулы предъявлялись не менее 50 раз и в случайном порядке.
Во второй серии испытуемый сам выполнял указанную деятельность пальцами левой руки, а в третьей серии — пальцами правой руки.
В четвертой серии («наблюдение за репродукцией ритма») испытуемый наблюдал за оператором, который вначале запоминал пятисекундный ритм, затем средним и указательным пальцами ведущей руки воспроизводил этот ритм, периодически нажимая на клавишу «пробел». Период ритма задавался зрительными стимулами (светлый квадрат, появляющийся на 200 мс в центре затемненного экрана монитора).
В пятой серии испытуемый сам выполнял указанную деятельность пальцами левой руки, а в шестой серии — пальцами правой руки.
Перед и в процессе выполнения деятельности регистрировали ЭЭГ с помощью электроэнцефалографа «Энцефалан-131-03» («Медиком МТД», Россия) в лобных, центральных, височных, теменных и затылочных отведениях по системе «10–20%». С целью исключения артефактов, связанных с движением глаз и мышечной активностью, проводили электроокулографию и электромиографию мышц шеи и лба. При вводе аналоговых сигналов в ЭВМ частота дискретизации составляла 250 Гц. С целью изучения корковых связей на частоте мю-ритма ЭЭГ предварительно фильтровали. Для этого применяли полосовой фильтр Баттерворта 20-го порядка с коэффициентом подавления частот выше 13 Гц не менее 80 дБ и частот ниже 8 Гц — не менее 40 дБ. При обработке полученных данных подсчитывали максимальные значения кросс-корреляционных функций и спектральной мощности ЭЭГ на коротких отрезках записи ЭЭГ (1–1,5 с): за 3 с до нажатия на клавишу (этап «фон»); за 1,5 с до нажатия на клавишу (этап «подготовка»); сразу после нажатия на клавишу (этап «выполнение действия»). Полученные значения коэффициентов корреляции и уровня спектральной мощности усредняли отдельно для каждого этапа деятельности, для каждой серии и по всем испытуемым. При подсчете кросс-корреляционных функций опирались на существующие рекомендации [10], что максимальный временной сдвиг должен быть не более одной десятой длины реализации, которая выбиралась более или равной десяти периодам мю-ритма (1–1,5 с). Для описания спектра мощности ЭЭГ применяли преобразование Фурье. Расчет спектра выполняли с аппроксимацией на целые гармоники (8, 9, 10, 11, 12, 13 Гц), что позволяло существенно упростить последующую статистическую обработку и сравнительный анализ результатов. При статистической обработке данных использовали пакет MatLab v. 6.5, непараметрический дисперсионный анализ и критерий Вилкоксона для связанных выборок.
В фМРТ-исследовании приняли участие 10 здоровых добровольцев — юношей в возрасте от 19 до 27 лет; средний возраст — 23 года. От каждого испытуемого было получено добровольное согласие на участие в эксперименте. Разрешение на проведение данных исследований было предоставлено Этической комиссией НИЦ «Курчатовский институт». В ходе предварительного обследования вышеуказанными методами исследовали особенности латеральной организации мозга с определением ведущей руки и речевого полушария. Результаты фМРТ получены в Курчатовском комплексе НБИКС-технологий НИЦ «Курчатовский институт» на томографе 3 Tл Magnetom Verio (Siemens, Германия).
Исследование включало несколько серий экспериментов. В первой серии «наблюдение за отмериванием интервала времени» испытуемый просматривал видеоролик с изображением руки оператора, пальцы которой паузой между двумя нажатиями на клавишу «пробел» отмеривали временной интервал 0,8 с.
Во второй серии «отмеривание интервала времени» испытуемый в процессе сканирования сам отмеривал данный временной интервал, нажимая на кнопку пальцами правой или левой руки в зависимости от инструкции.
В третьей серии «наблюдение за репродукцией ритма» испытуемому предварительно показывали видеоролик, на котором в центре экрана периодически (с интервалом 5 с) появлялся белый квадрат. После этого демонстрировали видео, на котором была показана рука оператора, воспроизводящая заданный ритм путем нажима средним и указательным пальцами на клавишу «пробел».
В четвертой серии «репродукция ритма» испытуемый сам воспроизводил заданный ритм, поочередно нажимая кнопки правой или левой рукой в зависимости от инструкции.
Все полученные фМРТ-данные были обработаны с помощью пакета программ SPM8. В рамках каждой из серий эксперимента были проведены попарные сравнения на основе статистики Стьюдента и получены индивидуальные и среднегрупповые статистические карты с уровнем значимости p<0,001. Все статистические карты наносили на шаблонное Т1-взвешенное изображение и производили анатомическую привязку «активных» вокселей к атласу ICN.
Результаты и обсуждение
Результаты исследования электрической активности мозга при наблюдении и восприятии времени. По данным анкетирования испытуемые распределились следующим образом: правшей — 27, амбидекстров — 4, левшей — 0. По результатам дихотического теста были выделены лица с левополушарным доминированием в отношении речи — 13; лица с правополушарным доминированием в отношении речи — 3; лица, у которых не выявлено доминантное в отношении речи полушарие, — 15.
Проведенные исследования позволили обнаружить статистически значимые изменения спектральной мощности ЭЭГ на частотах мю-ритма на разных этапах выполняемой деятельности. Оказалось, что характер этих изменений зависит от частоты данного ритма, латеральной организации мозга, вида и этапа выполняемой деятельности. Так, например, в серии с наблюдением за отмериванием длительности в отведениях С3 и С4 на частоте 8 Гц на этапе выполнения действия отмечается статистически значимое (p=0,034) по сравнению с фоном снижение спектральной мощности ЭЭГ. В серии с отмериванием длительности левой рукой на том же этапе выполнения действия и на той же частоте наблюдается статистически значимое (p=0,033) снижение спектральной мощности ЭЭГ в отведениях С4 и Сz. В серии с отмериванием длительности правой рукой на этом этапе в отведении Cz наблюдается статистически значимое (p=0,015) снижение спектральной мощности ЭЭГ на частоте 13 Гц. В серии «наблюдение за репродукцией ритма» на этапе «подготовка» по сравнению с «фоном» отмечается статистически значимое (p<0,05) снижение спектральной мощности ЭЭГ в отведении С3 на частотах 9 и 10 Гц, а в отведении С4 — на частоте 10 Гц. В то же время на этапе «выполнение действия» по сравнению с «фоном» статистически значимые изменения спектральной мощности ЭЭГ на частотах мю-ритма отсутствовали. По всей вероятности, обнаруженное на отдельных частотах снижение спектральной мощности мю-ритма на этапах «подготовка» и «выполнение действия» отражает активацию «двигательных» зеркальных нейронов [11].
Сравнение серий с репродукцией ритма левой и правой рукой позволило обнаружить межполушарные различия в значениях спектральной мощности ЭЭГ на частотах мю-ритма. Оказалось, что на этапе «выполнение действия» при репродукции ритма левой рукой спектральная мощность ЭЭГ на частоте 11 Гц в отведении С4 статистически значимо (p<0,05) ниже, чем в том же отведении при репродукции ритма правой рукой.
Дисперсионный анализ позволил обнаружить статистически значимое влияние факторов «ведущая рука» и «речевое полушарие» на спектральные характеристики мю-ритма в зависимости от этапа выполняемой деятельности. В частности, в серии наблюдений за отмериванием длительности на этапе «выполнение действия» на частотах 8 и 9 Гц в отведениях С3, С4 и Сz обнаружено статистически значимое (p=0,041–0,012) влияние фактора «ведущая рука» на спектральные характеристики мю-ритма. В серии c отмериванием длительности правой рукой обнаружено влияние фактора «речевое полушарие» на этапе «фон» в отведении С3 на частотах 8 и 9 Гц (p=0,004–0,002).
Анализ корковых взаимодействий на частоте мю-ритма при наблюдении и восприятии времени позволил обнаружить зависимость этих взаимодействий от этапа и вида выполняемой деятельности, а также от латеральной организации мозга. В частности, оказалось, что подготовка и выполнение действий, связанных с отмериванием длительности или репродукцией пятисекундного ритма, чаще всего сопровождаются статистически значимым (p<0,05) по сравнению с фоном усилением уровней корковых связей между центральными и лобными, височными, теменными и затылочными зонами коры. По всей вероятности, это усиление корковых связей отражает передачу сигналов из указанных зон в центральные области коры, где расположены «двигательные» зеркальные нейроны [11].
Зависимость корковых взаимодействий от этапа выполняемой деятельности при наблюдении и репродукции пятисекундного ритма иллюстрирует рис. 1.
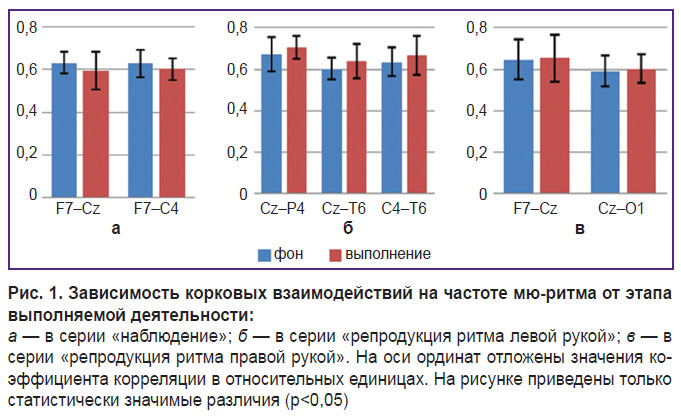 |
Рис. 1. Зависимость корковых взаимодействий на частоте мю-ритма от этапа выполняемой деятельности: а — в серии «наблюдение»; б — в серии «репродукция ритма левой рукой»; в — в серии «репродукция ритма правой рукой». На оси ординат отложены значения коэффициента корреляции в относительных единицах. На рисунке приведены только статистически значимые различия (p<0,05) |
Кроме того, изучение особенностей корковых взаимодействий при выполнении действий правой или левой рукой позволило обнаружить межполушарные различия уровней корковых связей. В частности, при репродукции ритма левой рукой (рис. 1, б) наблюдается статистически значимое усиление правополушарных связей, а при репродукции ритма правой рукой — усиление левополушарных связей (рис. 1, в).
Результаты фМРТ-исследования активности мозга при наблюдении и восприятии времени. Анализ данных фМРТ-сканирования мозга при наблюдении и восприятии времени позволил обнаружить существенные различия активации мозговых структур при наблюдении и выполнении действий, связанных с восприятием времени.
Результаты группового анализа фМРТ-данных по 4 сериям представлены на рис. 2–5.
|
Рис. 2. Полученные в рамках анализа серии 1 «наблюдение за отмериванием оператором интервала 0,8 с» групповые статистические карты, нанесенные на шаблонное Т1-взвешенное изображение высокого разрешения |
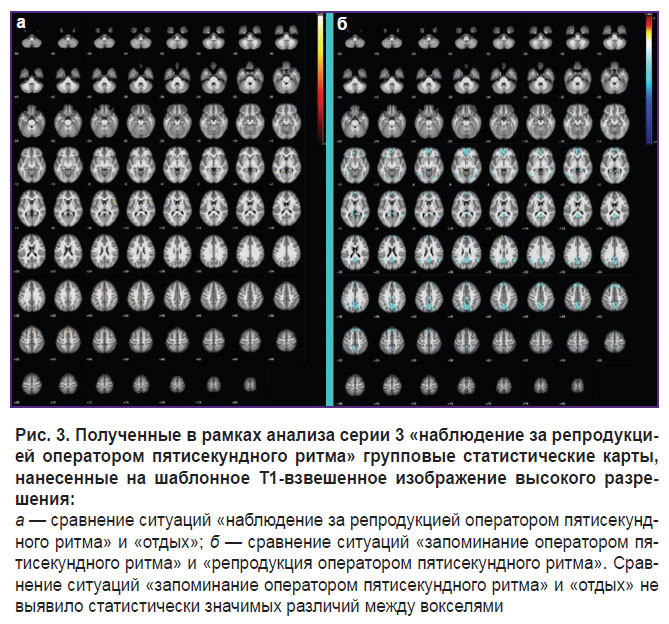
|
Рис. 3. Полученные в рамках анализа серии 3 «наблюдение за репродукцией оператором пятисекундного ритма» групповые статистические карты, нанесенные на шаблонное Т1-взвешенное изображение высокого разрешения:
а — сравнение ситуаций «наблюдение за репродукцией оператором пятисекундного ритма» и «отдых»; б — сравнение ситуаций «запоминание оператором пятисекундного ритма» и «репродукция оператором пятисекундного ритма». Сравнение ситуаций «запоминание оператором пятисекундного ритма» и «отдых» не выявило статистически значимых различий между вокселями |
|
Рис. 4. Полученные в рамках анализа серии 2 «отмеривание интервала 0,8 с» групповые статистические карты, нанесенные на шаблонное Т1-взвешенное изображение высокого разрешения: а — сравнение ситуаций «отмеривание интервала 0,8 с левой рукой» и «отдых»; б — сравнение ситуаций «отмеривание интервала 0,8 с правой рукой» и «отмеривание интервала 0,8 с левой рукой»; в — сравнение ситуаций «отмеривание интервала 0,8 с правой рукой» и «отдых» |
|
Рис. 5. Полученные в рамках анализа серии 4 «репродукция пятисекундного ритма» групповые статистические карты, нанесенные на шаблонное Т1-взвешенное изображение высокого разрешения:
а — сравнение ситуаций «репродукция ритма левой рукой» и «отдых»; б — сравнение ситуаций «репродукция ритма правой рукой» и «репродукция ритма левой рукой»; в — сравнение ситуаций «репродукция ритма правой рукой» и «отдых» |
В частности, анализ результатов, представленных на рис. 2, указывает на то, что в случае, когда испытуемый наблюдает за действиями другого человека, который отмеривает интервал времени 0,8 с, включаются зоны моторной и сенсомоторной коры, дополнительной моторной области, а также зоны скорлупы и хвостатого ядра. Как известно, перечисленные зоны отвечают в основном за двигательную активность и могут активироваться как часть двигательной системы зеркальных нейронов при наблюдении за моторными движениями другого человека [11].
При наблюдении за репродукцией пятисекундного ритма (см. рис. 3) дополнительно активируются области мозга, относящиеся к супрамаргинальной извилине. Активируются также височные отделы мозга и области префронтальной коры, в которых предположительно находятся «двигательные» зеркальные нейроны [11].
При отмеривании интервала времени 0,8 с и репродукции пятисекундного ритма (см. рис. 4 и 5) дополнительно активируются районы поясной извилины (что может быть связано с актуализацией памяти); области левого таламуса; височные отделы мозга; зрительная кора; области прекунеуса (предклинья), играющего важную роль в процессах самосознания, координации движений; и области мозжечка cerebellum crus 1 left и cerebellum 6 left, которые участвуют в процессах восприятия времени [12]. Полученные результаты свидетельствуют о том, что в данных процессах участвует целый комплекс мозговых структур. Условно их можно разделить на структуры, относящиеся к системе зеркальных нейронов, и структуры, относящиеся к системе отсчета времени, которые функционируют совместно. К первой системе, по-видимому, относятся зоны моторной и сенсомоторной коры, дополнительной моторной области, зоны скорлупы и хвостатого ядра, а также области мозга, принадлежащие супрамаргинальной извилине (supramarginal gyrus), которые активируются при наблюдении за репродукцией ритма и отмериванием интервалов времени. В пользу этого свидетельствуют и некоторые литературные данные [11] о том, что двигательная зеркальная система включает нейроны передней части нижнетеменной коры, нижней части прецентральной извилины, а также задней части нижнелобной извилины.
Ко второй системе, по-видимому, относятся зоны поясной извилины, области левого таламуса, височные отделы мозга, зрительная кора, области прекунеуса (предклинья) и области мозжечка (cerebellum crus 1 left и cerebellum 6 left), которые активируются при репродукции ритма и отмеривании интервалов времени. На это указывают и литературные данные, свидетельствующие о том, что важную роль в процессах восприятия времени играют мозжечок и базальные ганглии [12], гиппокамп [13], а также внутритеменная, фронтальная и слуховая зоны коры [14, 15].
Заключение
Наблюдение и выполнение деятельности, связанной с восприятием времени, сопровождается депрессией мю-ритма и вместе с тем усилением уровней корковых связей на частоте этого ритма. Характер этих изменений существенно зависит от латеральной организации мозга, частоты мю-ритма, вида и этапа выполняемой деятельности.
В процессах восприятия времени, по данным фМРТ-исследований, фактически участвуют две системы, которые работают совместно: 1) система зеркальных нейронов, включающая зоны моторной, сенсомоторной коры, дополнительной моторной области, а также зоны скорлупы, хвостатого ядра, области мозга, относящиеся к супрамаргинальной извилине, височные отделы мозга, области префронтальной коры и 2) система отсчета времени, включающая районы поясной извилины, области левого таламуса, височные отделы мозга, зрительную кору, области предклинья и области мозжечка.
Результаты исследования имеют важное теоретическое значение для понимания системных механизмов восприятия времени.
Финансирование исследования. Работа выполнена при финансовой поддержке гранта Российского фонда фундаментальных исследований №18-013-00758 и внутреннего проекта Национального исследовательского центра «Курчатовский институт» (приказ №1649 от 11 июля 2018 года).
Конфликт интересов отсутствует.
Литература
- Skoyles J.R. Gesture, language origins, and right handedness. Psycoloquy 2000; 11(24).
- Dapretto M., Davies M.S., Pfeifer J.H., Scott A.A., Sigman M., Bookheimer S.Y., Iacoboni M. Understanding emotions in others: mirror neuron dysfunction in children with autism spectrum disorders. Nat Neurosci 2005; 9(1): 28–30, https://doi.org/10.1038/nn1611.
- Аликина М.А., Махин С.А., Павленко В.Б. Амплитудно-частотные, топографические, возрастные особенности и функциональное значение сенсомоторного ритма ЭЭГ. Ученые записки Крымского федерального университета имени В.И. Вернадского. Биология. Химия 2016; 2(2): 3–24.
- Hobson H.M., Bishop D.V.M. Mu suppression — a good measure of the human mirror neuron system? Cortex 2016; 82: 290–310, https://doi.org/10.1016/j.cortex.2016.03.019.
- Pfurtscheller G., Neuper C., Krausz G. Functional dissociation of lower and upper frequency mu rhythms in relation to voluntary limb movement. Clin Neurophysiol 2000; 111(10): 1873–1879, https://doi.org/10.1016/s1388-2457(00)00428-4.
- Yang C.-Y., Decety J., Lee S., Chen C., Cheng Y. Gender differences in the mu rhythm during empathy for pain: an electroencephalographic study. Brain Res 2009; 1251: 176–184, https://doi.org/10.1016/j.brainres.2008.11.062.
- Anwar M.N., Navid M.S., Khan M., Kitajo K. A possible correlation between performance IQ, visuomotor adaptation ability and mu suppression. Brain Res 2015; 1603: 84–93, https://doi.org/10.1016/j.brainres.2015.01.045.
- Höller Y., Bergmann J., Kronbichler M., Crone J.S., Schmid E.V., Thomschewski A., Butz K., Schütze V., Höller P., Trinka E. Real movement vs. motor imagery in healthy subjects. Int J Psychophysiol 2013; 87(1): 35–41, https://doi.org/10.1016/j.ijpsycho.2012.10.015.
- Махин С.А., Макаричева А.А., Луцюк Н.В., Черный С.В., Орехова Л.С. Взаимосвязь между индивидуальным уровнем эмоционального интеллекта и реактивностью сенсомоторного ритма при синхронной имитации движений другого человека. Ученые записки Таврического национального университета имени В.И. Вернадского. Серия «Биология, химия» 2013; 26(4): 121–131.
- Bendat J.S., Piersol A.G. Random data: analysis and measurement procedures. Wiley; 2010.
- Rizzolatti G., Sinigaglia C. Mirrors in the brain: how our minds share actions and emotions. New York: Oxford University Press; 2008.
- Jueptner M., Rijntjes M., Weiller C., Faiss J.H., Timmann D., Mueller S.P., Diener H.C. Localization of a cerebellar timing process using PET. Neurology 1995; 45(8): 1540–1545, https://doi.org/10.1212/wnl.45.8.1540.
- Меринг Т.А. О различных формах отражения времени мозгом. Вопросы философии 1975; 7: 119–127.
- Leon M.I., Shadlen M.N. Representation of time by neurons in the posterior parietal cortex of the macaque. Neuron 2003; 38(2): 317–327, https://doi.org/10.1016/s0896-6273(03)00185-5.
- Сысоева О.В., Вартанов А.В. Отражение длительности стимула в характеристиках вызванного потенциала (часть 1). Психологический журнал 2004; 25(1): 101–110.








