Обоснование выбора оптимального стока крыс в качестве модельного объекта для исследований по поиску терапевтической мишени с целью коррекции расстройств полового поведения
Цель исследования — выявление механизмов, опосредующих различия в половом поведении самцов крыс Sprague Dawley и Wistar, для идентификации наиболее подходящего стока при исследованиях по поиску потенциальной терапевтической мишени для коррекции расстройств, связанных с мужским половым поведением.
Материалы и методы. Эксперименты проводили на половозрелых самцах крыс двух стоков (Sprague Dawley и Wistar) массой 350–450 г в возрасте от 3 до 6 мес. Особенности поведения животных исследовали с использованием стандартных тестов по изучению социального взаимодействия и рефлекторной активности, а также процедуры спаривания с рецептивными самками. С целью определения роли глицинергической нейропередачи в половом поведении применяли метод хронического билатерального вживления внутримозговых канюлей для осуществления серии последовательных фармакологических манипуляций по изменению уровня активности глициновых рецепторов в медиальной преоптической области (МПО) гипоталамуса исследуемых животных.
Результаты. Анализ полученных результатов выявил статистически значимые межстоковые различия на заключительной консуматорной стадии полового поведения как у интактных животных, так и после фармакологической активации глициновых рецепторов в МПО. Кроме того, в тесте «открытое поле» при исследовании груминга были обнаружены статистически значимые межстоковые различия, которые также присутствовали как у интактных животных, так и после фармакологической активации глициновых рецепторов в МПО, причем наблюдавшиеся различия исчезали после блокады глициновых рецепторов в МПО. В тесте Кроули при изучении социального взаимодействия статистически значимых различий между стоками не обнаружено.
Таким образом, отличия в половом поведении самцов крыс стоков Sprague Dawley и Wistar обусловлены разницей в уровне тревожности, что в свою очередь связано с различиями функционирования глицинергической нейропередачи в нейронных сетях МПО гипоталамуса у самцов крыс разных стоков.
Заключение. Для изучения связи уровня тревожности и полового поведения оптимальным является выбор стока крыс Wistar, поскольку половое поведение у самцов данного стока более чувствительно к стрессу, чем у самцов стока Sprague Dawley. В то же время для моделирования различных нарушений полового поведения, не связанных с тревожностью, предпочтение следует отдавать стоку Sprague Dawley, самцы которого обладают более стабильным половым поведением, менее зависящим от уровня тревожности.
Введение
Важной частью как фундаментальных, так и прикладных доклинических исследований новых лекарственных средств являются модельные организмы, среди которых наиболее популярны лабораторные грызуны, в частности инбредные линии и аутбредные стоки мышей и крыс [1–3]. Так, эксперименты по изучению различных аспектов полового поведения чаще всего проводят на крысах из аутбредных стоков Sprague Dawley и Wistar. Несмотря на важность выбора адекватной модели для доклинических исследований, существует всего несколько работ [4, 5], в которых рассматривается разница в половом поведении между крысами этих двух стоков. В ранее проведенных экспериментах по выявлению роли глициновой нейропередачи в медиальной преоптической области (МПО) гипоталамуса у стоков крыс Sprague Dawley [6] и Wistar [7] наша исследовательская группа обнаружила существенную разницу в половом поведении интактных самцов и самцов после фармакологической модуляции глициновых рецепторов в МПО.
Одними из факторов, обусловливающими различия, могут быть комплексная природа полового поведения, включающего в себя социальный и эмоциональный компоненты, а также вовлеченность МПО в контроль как социального поведения, так и уровня тревожности [8, 9]. МПО является важным звеном нейронных сетей мозга, которые участвуют в регуляции большинства рефлексов, связанных с половым поведением самцов различных видов позвоночных животных [10]. МПО гипоталамуса рассматривается в качестве структуры-кандидата, интегрирующего гомеостатическую информацию (в том числе об уровне стресса) с репродуктивным поведением, а также устанавливающего связь между нейроэндокринологическими факторами стресса и основными причинами половых расстройств, включая психические, сердечно-сосудистые и метаболические заболевания [11]. Именно МПО как структура головного мозга характеризуется высоким уровнем экспрессии глициновых рецепторов [12], нарушения в работе которых способны приводить к тяжелым неврологическим расстройствам, включая гиперэкплексию и эпилепсию [13]. Вероятно, глициновые рецепторы МПО могут являться потенциальной терапевтической мишенью для коррекции расстройств, связанных с мужским половым поведением.
Цель исследования — выявление механизмов, опосредующих различия в половом поведении самцов крыс Sprague Dawley и Wistar, для идентификации наиболее подходящего стока при исследованиях по поиску потенциальной терапевтической мишени для коррекции расстройств, связанных с мужским половым поведением.
Материалы и методы
Эксперименты проводили на половозрелых самцах и самках крыс стоков Sprague Dawley и Wistar в возрасте от 3 до 6 мес массой 350–450 г. Самки использовались только для тестирования полового поведения самцов. Исследование было одобрено Этическим комитетом Университета Умео (Швеция). Все экспериментальные и хирургические процедуры выполнены в соответствии с нормативами, указанными в руководстве «Guide for the Care and Use of Laboratory Animals (ILAR publication, 1996, National Academy Press)», межгосударственном стандарте ГОСТ 33216-2014 «Руководство по содержанию и уходу за лабораторными животными. Правила содержания и ухода за лабораторными грызунами и кроликами».
Порядок пре- и постоперативного тестирования поведения был идентичен для всех используемых поведенческих методик. Стандартная схема включала в себя однократное преоперативное тестирование интактных животных с последующей операцией по билатеральному вживлению канюль в MПO. После 7 дней восстановительного периода тестирование возобновляли. Всем животным в случайном порядке вводили раствор глицина — 1 мМ (агониста глициновых рецепторов), стрихнина — 20 мкМ (блокатора глициновых рецепторов в выбранной концентрации) и раствора Рингера для теплокровных в качестве контрольного. Интервал между тестами составлял 3 дня. После завершения экспериментов проводили гистологический контроль расположения кончиков канюль в мозговой ткани.
Регистрация поведенческих параметров при спаривании. Схема тестирования полового поведения самцов, хирургическая и фармакологическая подготовка рецептивных самок для спаривания, методика анализа параметров полового поведения самцов крыс, а также процедура вживления хронических билатеральных внутримозговых канюль и способ введения агонистов и антагонистов глициновых рецепторов в МПО крыс описаны нами ранее [6, 7]. Для оценки эффективности копуляции рассчитывали индекс интромиссий [14].
Регистрация поведенческих параметров при исследовании социального взаимодействия в тесте Кроули. Экспериментальная установка представляла собой прямоугольный бокс, изготовленный из прозрачного пластика размером 90×60×34 см и разделенный на 3 отсека 60×30 см (А, В, MID). Поведенческое тестирование состояло из 3 этапов длительностью по 5 мин каждый. С помощью SMART Video Tracking System v. 3.0 (Panlab, Испания) автоматически регистрировали время нахождения экспериментального животного в каждом из трех отсеков в соответствии с традиционным протоколом исследования.
Регистрация поведенческих параметров в тесте «открытое поле». Установка «открытое поле» (Panlab) представляла собой площадку 100×100 см, ограниченную непрозрачными стенками высотой 50 см. Поверхность установки была разделена на квадраты 20×20 см: 16 внешних квадратов, расположенных по периметру установки, и 9 внутренних квадратов. В ходе тестирования регистрировали частоту и длительность груминга, а также паттерн последовательных движений при груминге, который позволил выделить эффект «незавершенного груминга».
Обработка результатов. Статистический анализ данных, полученных в ходе поведенческих экспериментов, осуществляли с использованием программного обеспечения Prism Windows 5 (GraphPad, США). Анализ различий между группами проводили с помощью непараметрического U-критерия Манна–Уитни, поскольку полученные данные имели распределение, отличное от нормального. Проверку распределения данных на предмет соответствия нормальному закону выполняли по критерию Шапиро–Уилка. Критическое значение уровня статистической значимости при проверке нулевых гипотез принималось равным 0,05. Результаты представляли в виде медианы и процентилей — Me [25; 75].
Результаты
Интактные самцы
Анализ полового поведения. Сравнение аппетивной фазы полового поведения по параметру «длительность латентного периода интромиссии» не выявило статистически значимых различий (p=0,38) у интактных самцов стока Sprague Dawley (119,0 [44,5; 212,5] c; n=9) и стока Wistar (69,0 [26,0; 173,0] с; n=11). В силу того, что самцы крыс стока Wistar почти не совершали маунтингов как во время аппетивной, так и во время консуматорной фазы полового поведения, статистическое сравнение животных двух стоков по параметру «длительность латентного периода маунтинга» не представлялось возможным. При этом длительность данного параметра у крыс стока Sprague Dawley составляла 91,0 [45,5; 120,5] с.
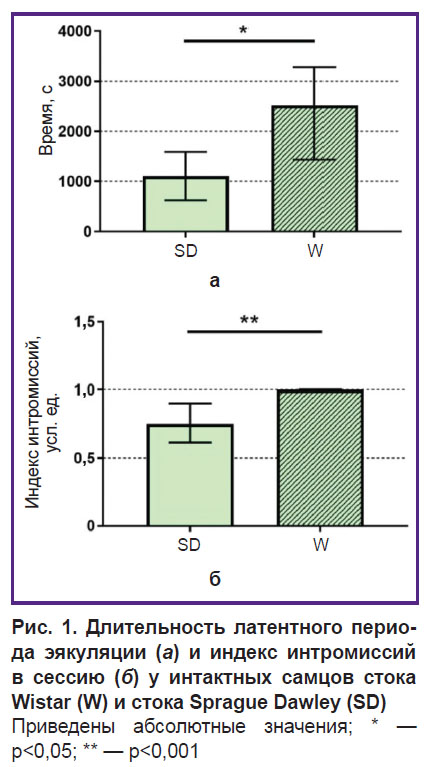
Консуматорная фаза полового поведения у интактных самцов стоков Sprague Dawley и Wistar статистически значимо (p<0,05) различалась по таким параметрам, как длительность латентного периода эякуляции и индекс интромиссий в сессию. Так, длительность латентного периода эякуляции была в 2 раза выше у самцов стока Wistar (2517,0 [1435,0; 3282,0] c; n=11) по сравнению со стоком Sprague Dawley (1102,0 [624,5; 1589,0] с; n=9), p=0,01 (рис. 1, а). В связи с тем, что самцы стока Wistar почти не совершали маунтингов, индекс интромиссий в сессию у них был выше (1,0 [1,0; 1,0] усл. ед.; n=11), чем у самцов стока Sprague Dawley (0,75 [0,61; 0,90] усл. ед.; n=9), p=0,0002 (рис. 1, б).
|
Рис. 1. Длительность латентного периода эякуляции (а) и индекс интромиссий в сессию (б) у интактных самцов стока Wistar (W) и стока Sprague Dawley (SD) Приведены абсолютные значения; * — p<0,05; ** — p<0,001 |
Таким образом, результаты экспериментов показывают существенные различия в половом поведении самцов двух стоков. Крысам стока Sprague Dawley требуется меньше времени для достижения эякуляции, что свидетельствует о большей эффективности спаривания у самцов этого стока по сравнению с самцами стока Wistar [15].
Анализ социального взаимодействия. На первом этапе тестирования, при отсутствии других животных внутри экспериментальной установки, самцы обоих стоков проводили во всех 3 отсеках (А, В, MID) примерно одинаковое количество времени. На втором этапе эксперимента (отсек А — резидент, отсек В — пусто) самцы обоих стоков проводили больше времени в отсеке с резидентом. На третьем этапе эксперимента (отсек А — резидент, отсек В — интрудер) самцы обоих стоков проводили больше времени в отсеке с интрудером. Такое поведение соответствует норме [16].
Таким образом, сходство социальной активности самцов стоков Sprague Dawley и Wistar в тесте Кроули свидетельствует о том, что социальное поведение не является фактором, опосредующим различия в половом поведении самцов этих двух стоков.
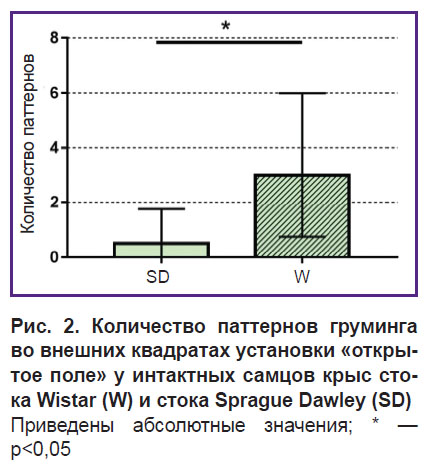
Анализ степени тревожности. При анализе поведенческих параметров обнаружены статистически значимые различия уровня тревожности. Так, количество паттернов груминга во внешних квадратах установки «открытое поле» было выше у самцов крыс стока Wistar (3,0 [0,75; 6,0]; n=10) по сравнению с самцами стока Sprague Dawley (0,5 [0; 1,75]; n=12), p=0,03 (рис. 2). При этом процедура груминга у крыс Wistar характеризовалась как «незавершенный груминг».
|
Рис. 2. Количество паттернов груминга во внешних квадратах установки «открытое поле» у интактных самцов крыс стока Wistar (W) и стока Sprague Dawley (SD) Приведены абсолютные значения; * — p<0,05 |
Полученные результаты тестирования свидетельствуют о том, что различия в половом поведении самцов двух стоков обусловлены более высоким уровнем тревожности крыс стока Wistar.
Самцы после билатеральных микроинъекций растворов глицина и стрихнина в медиальной преоптической области
Анализ полового поведения. После стимуляции процессов торможения в нейронных сетях путем билатеральных микроинъекций 1 мМ глицина у 4 из 8 самцов стока Wistar полностью (отсутствие эякуляции) либо частично исчезало половое поведение, в то время как самцы Sprague Dawley сохраняли способность к копуляции в 100% случаев. Более того, длительность латентного периода эякуляции после микроинъекций 1 мМ глицина у самцов стока Sprague Dawley снижалась (887,5 [556,0; 1063,0] с), что свидетельствует о повышении эффективности копуляции самцов [15]. Ввиду малого размера выборки самцов Wistar, совершивших полноценный копуляторный акт после микроинъекций 1 мМ глицина (n=4), статистическое сравнение параметров полового поведения самцов двух стоков проводили только на следующем этапе.
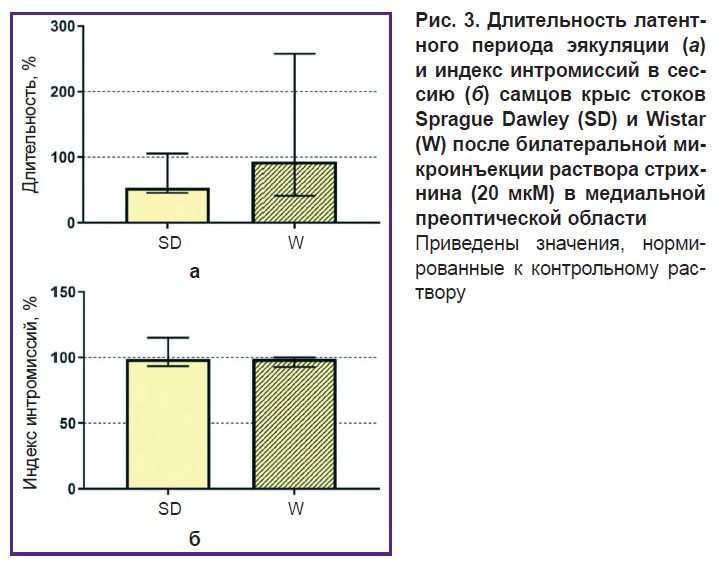
После билатеральной микроинъекции 20 мкМ стрихнина у самцов стока Wistar укорачивался латентный период эякуляции (1905,0 [994,5; 3078,0] с) в сессию по сравнению с большой задержкой или полным отсутствием эякуляции при стимуляции торможения, благодаря чему статистически значимая разница между двумя стоками по этому параметру исчезала (рис. 3, а).
|
Рис. 3. Длительность латентного периода эякуляции (а) и индекс интромиссий в сессию (б) самцов крыс стоков Sprague Dawley (SD) и Wistar (W) после билатеральной микроинъекции раствора стрихнина (20 мкМ) в медиальной преоптической области Приведены значения, нормированные к контрольному раствору |
Кроме того, микроинъекции 20 мкМ стрихнина приводили к появлению паттернов маунтинга у самцов стока Wistar, что в свою очередь влекло за собой исчезновение статистически значимых различий между самцами двух стоков по параметру «индекс интромиссий» (рис. 3, б). В то же время микроинъекции 20 мкМ стрихнина практически не влияли на изменение длительности латентного периода эякуляции у самцов стока Sprague Dawley (1044,0 [536,5; 1497,0] с) относительно интактных значений.
Таким образом, активация глицинергического торможения в нейронных сетях МПО гипоталамуса способствовала полному или частичному исчезновению полового поведения у самцов стока Wistar и, наоборот, увеличению эффективности копуляции у самцов стока Sprague Dawley, а ингибирование глицинергического торможения стрихнином приводило к восстановлению эффективности копуляции самцов стока Wistar и не изменяло параметры полового поведения самцов стока Sprague Dawley.
Анализ социальной активности. Статистически значимых различий в социальной активности (тест Кроули) у самцов крыс стоков Sprague Dawley и Wistar после билатеральных микроинъекций глицина и стрихнина не выявлено.
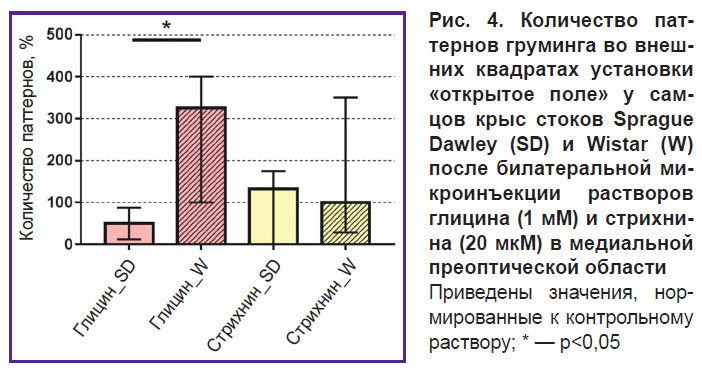
Анализ степени тревожности. Фармакологическая манипуляция глицинергической синаптической передачи в нейронных сетях МПО с помощью микроинъекций 1 мМ глицина приводила к увеличению количества «незавершенных паттернов груминга» во внешних квадратах установки «открытое поле» у самцов стока Wistar (5,0 [3,0; 8,5]), что свидетельствовало об усилении тревожности [17]. Кроме того, после билатеральной микроинъекции глицина сохранялась статистически значимая разница между количеством паттернов груминга у самцов стока Wistar и самцов стока Sprague Dawley (p=0,02) (рис. 4). Однако последующие микроинъекции 20 мкМ стрихнина приводили к снижению количества паттернов груминга у самцов стока Wistar (3,50 [1,75; 6,0]) по сравнению с уровнем при стимуляции торможения в нейронных сетях, в связи с чем статистическая значимость различий между количеством паттернов груминга у самцов двух стоков исчезала (см. рис. 4).
|
Рис. 4. Количество паттернов груминга во внешних квадратах установки «открытое поле» у самцов крыс стоков Sprague Dawley (SD) и Wistar (W) после билатеральной микроинъекции растворов глицина (1 мМ) и стрихнина (20 мкМ) в медиальной преоптической области Приведены значения, нормированные к контрольному раствору; * — p<0,05 |
Таким образом, билатеральное введение глицина/стрихнина выявило различия в ответной реакции поведения крыс двух стоков на стимуляцию/ингибирование глицинергической передачи в МПО гипоталамуса при прохождении теста «открытое поле».
Обсуждение
Анализ полученных результатов тестирования полового поведения интактных животных выявил статистически значимые различия (p<0,05) в консуматорном половом поведении самцов Sprague Dawley и Wistar. Наиболее заметным отличием оказалось почти полное отсутствие паттерна «маунтинг» у самцов стока Wistar. В литературе есть упоминание о том, что самцы крыс стока Wistar совершают меньше маунтингов и больше интромиссий, чем самцы стока Sprague Dawley [4]. Кроме того, самцам стока Wistar требовалось больше времени и интромиссий для достижения эякуляции. Данное различие может свидетельствовать о меньшей эффективности копуляции самцов стока Wistar. Возможно, это связано с более высоким уровнем тревожности у крыс данного стока, так как, согласно модели «воронка» [18], тревожные самцы могут тратить больше времени на проверку безопасности окружающей среды, тем самым снижая эффективность копуляции.
Тест «открытое поле» подтвердил наличие более высокой степени тревожности у интактных самцов стока Wistar. Количество груминга у этих животных было выше, чем у самцов стока Sprague Dawley. Причем особенностью груминга у крыс стока Wistar являлась его «незавершенность», свидетельствующая о более тревожном состоянии животного [17, 19].
Выявленные различия в половом поведении самцов стоков Sprague Dawley и Wistar, вероятно, обусловлены различием в структуре тормозных процессов в нейронных сетях МПО гипоталамуса. Анализ полового поведения после билатеральных микроинъекций растворов глицина и стрихнина в МПО выявил различия между самцами данных стоков. Микроинъекции глицина в МПО у самцов стока Wistar приводили к полной или частичной неспособности копулировать, в то время как эти же микроинъекции у самцов Sprague Dawley, наоборот, повышали эффективность копуляции, о чем свидетельствовало снижение длительности латентного периода эякуляции [15].
Известно, что глицин может взаимодействовать не только с собственным специфическим рецептором, оказывающим тормозное воздействие на нейроны [20], но и с регуляторным сайтом ионотропного рецептора глутамата, селективно связывающим N-метил-D-аспартат (NMDA-рецепторы) [21]. Следовательно, принимая во внимание тот факт, что микроинъекции глицина приводили к исчезновению копуляции у самцов стока Wistar и, наоборот, способствовали увеличению эффективности копуляции у самцов стока Sprague Dawley, можно предположить, что животные этих двух стоков имеют разное соотношение глициновых и NMDA-рецепторов. В пользу данной гипотезы свидетельствует появление паттерна «маунтинг» у самцов стока Wistar после микроинъекций стрихнина в МПО. Поскольку маунтинги рассматриваются в качестве самостоятельного паттерна при копуляции [15], их возникновение можно расценивать как обогащение поведенческого репертуара самцов. В то же время микроинъекции стрихнина в МПО не приводят к значимым изменениям в поведении самцов стока Sprague Dawley.
Участие глицинергической нейропередачи МПО в регуляции полового поведения самцов стоков Sprague Dawley и Wistar может также опосредоваться через регуляцию степени тревожности животных. Данное предположение подтверждают результаты анализа теста «открытое поле» при фармакологической манипуляции глицинергической нейропередачи в МПО крыс обоих стоков. Микроинъекции глицина увеличивают количество паттернов «незавершенного» груминга у самцов стока Wistar, а инъекции стрихнина приводят к снижению выраженности данной реакции.
В то же время анализ социального взаимодействия (тест Кроули) интактных животных и животных с активацией/ингибированием глицинергической передачи не выявил разницы в поведении самцов стоков Sprague Dawley и Wistar, следовательно, социальная активность не является причиной различий в половом поведении самцов данных стоков.
Таким образом, обнаруженная в исследовании статистически значимая разница в половом поведении самцов крыс стоков Sprague Dawley и Wistar может быть обусловлена разным соотношением глициновых и NMDA-рецепторов в МПО гипоталамуса и связана с различной степенью тревожности у данных стоков, которая в свою очередь, вероятно, также вызвана различием в функционировании глицинергической передачи в МПО гипоталамуса. Данный механизм следует учитывать при выборе модельного стока крыс для исследования полового поведения в эксперименте.
Также представляется интересным дальнейшее изучение генетически детерминированных различий в экспрессии глициновых рецепторов МПО для более детального выявления роли глицинергической передачи МПО в регуляции индивидуального полового поведения.
Заключение
Для изучения связи уровня тревожности и полового поведения оптимальным является выбор стока крыс Wistar, поскольку половое поведение у самцов данного стока более чувствительно к стрессу, чем у самцов стока Sprague Dawley. В то же время для моделирования различных нарушений полового поведения, не связанных с тревожностью, предпочтение следует отдавать стоку Sprague Dawley, самцы которого обладают более стабильным половым поведением, менее зависящим от уровня тревожности.
Финансирование. Исследование не финансировалось никакими источниками.
Конфликт интересов. Авторы заявляют об отсутствии конфликта интересов.
Литература
- Mehta N., Sikka S., Rajasekaran M. Rat as an animal model for male erectile function evaluation in sexual medicine research. J Sex Med 2008; 5(6): 1278–1283, https://doi.org/10.1111/j.1743-6109.2008.00854.x.
- Skayneh H., Jishi B., Hleihel R., Hamieh M., Darwiche N., Bazarbachi A., El Sabban M., El Hajj H. A critical review of animal models used in acute myeloid leukemia pathophysiology. Genes (Basel) 2019; 10(8): 614, https://doi.org/10.3390/genes10080614.
- Manns M., Basbasse Y.E., Freund N., Ocklenburg S. Paw preferences in mice and rats: meta-analysis. Neurosci Biobehav Rev 2021; 127: 593–606, https://doi.org/10.1016/j.neubiorev.2021.05.011.
- McLean J.H., Dupeire W.A., Elder S.T. Strain differences in the mating behavior of Sprague-Dawley, Long-Evans, and Wistar male rats. Psychon Sci 1972; 175–176, https://doi.org/10.3758/bf03342585.
- Emery D.E., Larsson K. Rat strain differences in copulatory behavior after para-chlorophenylalanine and hormone treatment. J Comp Physiol Psychol 1979; 93(6): 1067–1084, https://doi.org/10.1037/h0077640.
- Zhuravleva Z.D., Lebedeva A.V., Volnova A.B., Mukhina I.V., Druzin M.Ya. The effect of glycine microinjections in the medial preoptic area of the hypothalamus on the sexual behavior of male rats. Neurochem J 2015; 9: 141–145, https://doi.org/10.1134/s1819712415020117.
- Zhuravleva Z.D., Mogutina M.A., Mukhina I.V., Druzin M.Ya. Effects of glycine receptors of the medial preoptic nucleus on sexual behavior of male Wistar rats. Bull Exp Biol Med 2020; 169(3): 299–301, https://doi.org/10.1007/s10517-020-04873-y.
- Zhao C., Chang L., Auger A.P., Gammie S.C., Riters L.V. Mu opioid receptors in the medial preoptic area govern social play behavior in adolescent male rats. Genes Brain Behav 2020; 19(7): e12662, https://doi.org/10.1111/gbb.12662.
- León Rodríguez D.A., Dueñas Z. Maternal separation during breastfeeding induces gender-dependent changes in anxiety and the GABA-A receptor alpha-subunit in adult Wistar rats. PLoS One 2013; 8(6): e68010, https://doi.org/10.1371/journal.pone.0068010.
- Hull E.M., Dominguez J.M. Neuroendocrine regulation of male sexual behavior. Compr Physiol 2019; 9(4): 1383–1410, https://doi.org/10.1002/cphy.c180018.
- Viau V. Functional cross-talk between the hypothalamic-pituitary-gonadal and -adrenal axes. J Neuroendocrinol 2002; 14(6): 506–513, https://doi.org/10.1046/j.1365-2826.2002.00798.x.
- Karlsson U., Druzin M., Johansson S. Cl– concentration changes and desensitization of GABAA and glycine receptors. J Gen Physiol 2011; 138(6): 609–626, https://doi.org/10.1085/jgp.201110674.
- Schaefer N., Roemer V., Janzen D., Villmann C. Impaired glycine receptor trafficking in neurological diseases. Front Mol Neurosci 2018; 11: 291, https://doi.org/10.3389/fnmol.2018.00291.
- Ågmo A. Male rat sexual behavior. Brain Res Brain Res Protoc 1997; 1(2): 203–209, https://doi.org/10.1016/s1385-299x(96)00036-0.
- Heijkoop R., Huijgens P.T., Snoeren E.M.S. Assessment of sexual behavior in rats: the potentials and pitfalls. Behav Brain Res 2018; 352: 70–80, https://doi.org/10.1016/j.bbr.2017.10.029.
- Kaidanovich-Beilin O., Lipina T., Vukobradovic I., Roder J., Woodgett J.R. Assessment of social interaction behaviors. J Vis Exp 2011; 48: 2473, https://doi.org/10.3791/2473.
- Kalueff A.V., Stewart A.M., Song C., Berridge K.C., Graybiel A.M., Fentress J.C. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat Rev Neurosci 2016; 17(1): 45–59, https://doi.org/10.1038/nrn.2015.8.
- Veening J.G., Coolen L.M. Neural mechanisms of sexual behavior in the male rat: emphasis on ejaculation-related circuits. Pharmacol Biochem Behav 2014; 121: 170–183, https://doi.org/10.1016/j.pbb.2013.12.017.
- Umryukhin P.E., Grigorchuk O.S. Behavior of rats in an open field test as a prognostic indicator of corticosterone levels before and after stress. Neurosci Behav Physi 2017; 47: 456–458, https://doi.org/10.1007/s11055-017-0421-3.
- Kumar A., Basak S., Rao S., Gicheru Y., Mayer M.L., Sansom M.S.P., Chakrapani S. Mechanisms of activation and desensitization of full-length glycine receptor in lipid nanodiscs. Nat Commun 2020; 11(1): 3752, https://doi.org/10.1038/s41467-020-17364-5.
- Yu A., Lau A.Y. Glutamate and glycine binding to the NMDA receptor. Structure 2018; 26(7): 1035–1043.e2, https://doi.org/10.1016/j.str.2018.05.004.








