Вирусные векторы для доставки генетического материала в клетку и их использование в нейробиологии (обзор)
Вирусные векторы — современный инструмент для доставки генетического материала в клетку. Рассмотрены различные типы вирусных векторов, таких как ретровирусные, аденоассоциированные и лентивирусные векторные системы, векторные системы на основе аденовирусов, вируса герпеса простого и поксвирусов. Более подробно представлены аденоассоциированные векторные системы. Их основные преимущества (способность интегрировать целевой ген в нужное место генома хозяина, что предотвращает нежелательные мутации; встраивание как в делящиеся, так и в покоящиеся клетки; широкий профиль трансдукции; низкий иммунный ответ; сильная и устойчивая экспрессия трансгена) сделали эти векторы популярным и универсальным инструментом для доставки генов in vitro и in vivo. Показаны возможности применения вирусных векторов в нейробиологии.
В настоящее время вирусные векторы — распространенный инструмент для доставки генетического материала в клетку. Именно благодаря особенностям жизненного цикла вирусов первые векторы (носители трансгенов) стали разрабатывать на их основе [1]. Вирусы переносят чужеродные гены, которые способны экспрессироваться в зараженных клетках [2]. Сейчас вирусы эволюционировали в специализированные молекулярные механизмы, способные эффективно транспортировать их геномы внутрь клеток, которые они заражают [3].
Разнообразие вирусных векторов велико, и каждый имеет свои преимущества и недостатки. Сейчас значительные усилия направляются на разработку вирусных векторов с улучшенными характеристиками безопасности и большей эффективностью доставки нуклеиновой кислоты в клетки, а также на обеспечение длительной и тканеспецифичной экспрессии введенного генетического материала. Потенциально вирусные векторы могут применяться как один из способов доставки генов в генной терапии новообразований и наследственных заболеваний.
Рекомбинантные аденоассоциированные вирусы (рААВ) являются одними из наиболее перспективных векторов доставки для генной терапии благодаря своим непатогенным свойствам, отсутствию иммуногенности со стороны хозяина и тропности к большинству клеток и тканей [4]. Аденоассоциированные вирусные векторы по своим характеристикам приближаются к идеальному вектору [5].
Вирусные векторы нашли свое применение и в нейробиологии в качестве систем эффективной доставки генов в нейроны и другие нейрональные клетки in vitro и in vivo. Вирусные векторы позволяют вести наблюдение за нейробиологическими функциями, изменять экспрессию целевых генов, метить клетки для определения их судьбы и менять физиологическое состояние специфических клеточных популяций. Использование вирусных векторов для доставки генов в нервную систему имеет большие перспективы как для фундаментальных исследований, так и для терапевтического применения.
Типы вирусных векторов, используемых в нейробиологии
Существуют вирусные и невирусные системы доставки генетического материала. Вирусные системы включают векторы, разработанные на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, лентивирусов и вирусов простого герпеса. Невирусные системы включают «голую» ДНК и липиды или полиэтилгликоль [4]. Независимо от того, какой геном имеют вирусные векторы в своем составе — ДНК или РНК, у них похожий жизненный цикл, который начинается с взаимодействия со специфическим рецептором на поверхности клетки. После адсорбции вириона на клеточной поверхности в процессе проникновения геном вирусов подвергается «раздеванию». При реализации внутриклеточной стадии жизненного цикла вирус осуществляет три молекулярных процесса: репликацию геномной нуклеиновой кислоты, транскрипцию и трансляцию. На каждой стадии вирус вмешивается в клеточные синтетические механизмы и подчиняет их своим задачам, создавая приоритеты для вирусных нуклеиновых кислот [6].
Ни ДНК, ни РНК не могут быть использованы в «голом» виде для доставки генов в клетки-мишени. Сахарофосфатный остов молекул нуклеиновых кислот располагается по их периферии полярными группами наружу, придавая им анионные свойства [7]. При физиологических значениях рН нуклеиновая кислота несет отрицательный заряд, отталкивающий ее от отрицательно заряженной наружной поверхности клеточной мембраны. Еще одно ограничение при проникновении в клетку нуклеиновой кислоте создает ее гидрофильность [8]. Все ее гидрофобные основания «повернуты» вовнутрь молекулы, поэтому она не может проникнуть через гидрофобный барьер клетки-мишени [9]. В сыворотке крови нуклеиновая кислота быстро деградирует под воздействием нуклеаз. Период полужизни немодифицированной интерферирующей РНК в сыворотке крови укладывается в 5–60 мин, для ДНК он составляет не более 10 мин [10]. Кроме того, нуклеиновые кислоты не способны специфически узнавать клетки-мишени. Поэтому для доставки генов в эукариотические клетки с начала 1980-х гг. разрабатываются векторные генетические конструкции [11].
Ретровирусные векторные системы
Ретровирусы относятся к группе вирусов, РНК-геном которых в инфицированных клетках конвертируется в ДНК. Геном ретровирусов образован плюс-цепью РНК и включает три структурных гена (gag, pol и env), фланкированных длинными терминальными повторами (LTR, long terminal repeat) [12]. В LTR содержатся регуляторные элементы, которые играют роль во встраивании ретровируса и необходимы для интеграции ДНК-копии генома вируса с геномом хозяина, а также определяют начало и конец вирусного генома. LTR также контролируют экспрессию вирусных генов [13]. Оболочка ретровирусов состоит из цитоплазматической мембраны зараженной клетки и вирусных белков [14]. Особенностью жизненного цикла ретровирусов является обратная транскрипция в зараженной клетке на матрице вирусной РНК с образованием двухцепочечной ДНК (провирус), которая затем встраивается в клеточный геном. Вирус встраивается в клеточный геном случайным образом, поэтому возрастает риск инсерционного мутагенеза. Так как эти вирусы заражают только делящиеся клетки (для проникновения в ядро необходимо разрушение ядерной оболочки клетки, происходящее в ходе митоза), ретровирусные векторы используют в основном для трансфекции клеток ex vivo или для лечения злокачественных опухолей [15]. Сами ретровирусные векторы получают на основе провируса, из которого удаляются гены gag, pol и env для предотвращения репродукции вируса и освобождения места для желаемого генетического материала. В вектор на основе ретровируса может быть включено до 8 тыс. пар оснований ДНК-вставки. Для репродукции вируса удаленные вирусные гены (gag, pol, env), которые кодируют вирусные белки и обеспечивают репродукцию вируса, встраивают в геном упаковывающей клеточной линии на разные хромосомы так, чтобы снизить вероятность обратной рекомбинации вирусных генов в исходный вирусный геном и образования вирусов, которые могут репродуцироваться [16].
Лентивирусные векторные системы
Лентивирусы относятся к семейству ретровирусов и в отличие от других ретровирусов инфицируют не только делящиеся, но и неделящиеся клетки. Наиболее исследованным лентивирусом является вирус иммунодефицита человека (ВИЧ). В связи со способностью лентивирусов включать большое количество генетического материала (до 8 тыс. пар оснований) и инфицировать делящиеся и неделящиеся клетки, эти вирусы являются перспективным вектором для доставки генов в условиях in vivo [17]. Геном ВИЧ включает три гена структурных белков (gag, pol и env) и 6 генов регуляторных белков (tat, rev, vpr, vpu, nef и vif) [18, 19]. Некоторые гены могут быть исключены из генома вируса и при этом не вызвать снижения вирусной способности к размножению и инфицированию клеток. Лентивирусный жизненный цикл похож на жизненные циклы других ретровирусов, за исключением способности заражать и неделящиеся клетки. Эта способность обеспечивается взаимодействием вирусного преинтеграционного комплекса с ядерной оболочкой и транспортом через нее [20]. Сборка лентивирусного вектора происходит в упаковывающих клетках — это перевиваемые клетки, осуществляющие синтез вирусоспецифических белков [21]. Упаковывающие клетки включают в себя пакующую, векторную и оболочечную кассеты, которые вместе позволяют собрать функциональную вирусную частицу [22, 23]. При этом их одновременная экспрессия не вызывает образования ретровирусных частиц, способных инициировать инфекционный процесс у человека [24].
Лентивирусы имеют сравнительно небольшой объем вставки целевого гена (до 8 тыс. пар оснований), могут обеспечить длительную экспрессию трансгена и индуцируют минимальный иммунный ответ организма-хозяина [25]. Некоторые авторы считают эти векторы малоподходящими для доставки in vivo, так как они увеличивают риск инсерционного мутагенеза. Однако при использовании лентивирусов для трансдукции дифференцированных клеток риск инсерционного мутагенеза меньше, чем при использовании других ретровирусов [26].
Векторные системы на основе аденовирусов
Аденовирусы — семейство ДНК-вирусов, несущих в своем составе одну двуцепочечную молекулу ДНК и лишенных липидной оболочки. Аденовирусы разделены на основе связывания со специфическими сыворотками на 51 серотип [27], а по способности агглютинировать эритроциты у людей, кроликов и мышей и по онкогенности для грызунов — еще на 6 субгрупп (от A до F) [28]. Аденовирусы разных субгрупп поражают различные органы и ткани человека [29].
Создание рекомбинантных аденовирусных векторов во многом схоже с созданием лентивирусных векторов. Аденовирусы, дефектные по репликации, получали за счет замены гена Е1, необходимого для репликации, на ген интереса, промотор и энхансер [30]. При этом такие рекомбинантные векторы эффективно размножаются в пакующих клетках, экспрессирующих продукт гена Е1. Неспособные к репликации вне пакующих клеток аденовирусные векторы используют для введения трансгенов в условиях in vivo [31]. Рекомбинантные аденовирусные векторы обеспечивают очень высокую экспрессию клонированных генов, но на короткое время (5–10 сут) — из-за иммунного ответа организма-реципиента [32]. Для решения этой проблемы было создано второе поколение аденовирусных векторов, у которых дополнительно к гену Е1 удалили гены, отвечающие за репликацию вируса, и оставили только элементы, определяющие начало и конец генома и вирусную пакующую последовательность [33]. Такие векторы способны к более длительной экспрессии генов [14].
Аденовирусы способны инфицировать большинство типов клеток (делящиеся и неделящиеся). Объем для вставки целевого гена составляет 20 тыс. пар оснований, что является достаточно большой емкостью рекомбинантных аденовирусных векторов. Аденовирусы реплицируются в ядре инфицированной клетки как эписомные элементы и обладают высокой эффективностью трансдукции. Так, например, после прямой внутричерепной инъекции рекомбинантного аденовируса была обнаружена его способность заражать нейроны, астроциты, олигодендроглию, эпендимоциты, хориоидальный эпителий и микроглию [34, 35].
Векторные системы на основе вируса простого герпеса
Вирусные векторы на основе вируса простого герпеса (herpes simplex virus, HSV) имеют конструкцию проще, чем векторы на основе аденовирусов. Сам вирус включает около 80 генов, один из которых (IE3) чаще всего замещается при создании вектора [36]. Исключены могут быть и другие гены, что позволяет увеличить объем вектора или заклонировать несколько генов интереса. Недостатками векторов на основе вирусов простого герпеса являются кратковременная экспрессия клонированных генов, токсичность для клеток-мишеней, низкая эффективность трансдукции и способность заражать только неделящиеся клетки [37].
Для создания вирусных векторов сейчас активно используются ампликоны — многократно повторяющиеся последовательности вируса простого герпеса, которые включают мономерные последовательности, организованные как конкатемеры [38]. Каждый мономер включает хотя бы один участок начала репликации вирусной ДНК (oriS или oriL) и последовательность для упаковки ДНК в вирусную частицу (pac) [39]. Молекулярное клонирование этих последовательностей в бактериальную плазмиду позволяет получить вектор, который упаковывает ДНК в вирион HSV. Такие векторные системы способны включать до 150 тыс. пар оснований чужеродной ДНК, что предоставляет возможность одним вектором доставлять в клетку-мишень несколько транскрипционных единиц, не вызывая при этом иммунного ответа и цитопатического эффекта [40]. Особенность жизненного цикла вирусов простого герпеса — наличие двух стадий инфицирования: активной (проникновение и репродукция в клетках эпителия, приводящие к лизису клеток) и латентной (внедрение и репродукция в нервных клетках) [41]. Лизис клеток происходит за счет того, что при репликации вируса сформировавшиеся вирионы покидают клетку путем почкования ядерной мембраны.
Вирусы простого герпеса — нейротрофичны и высокоэффективны при изучении ретроградного и антероградного транспорта в ЦНС, могут быть введены в неопасном латентном состоянии. HSV-векторы имеют большую генетическую емкость и могут обеспечить долгосрочную экспрессию трансгена, однако, как отмечалось выше, основным недостатком их являются токсичность для клеток и низкая эффективность трансдукции [24].
Векторные системы на основе поксвирусов
Поксвирусы — крупные вирусы, которые содержат двунитевую ДНК. Поксвирусный вектор позволяет включить до 25 тыс. пар нуклеотидов ДНК интереса без исключения генов самого вируса. Векторные системы на основе поксвирусов не имеют такого широкого применения, поскольку эукариотические промоторы неэффективно распознаются транскрипционными механизмами поксвирусов и для эффективной экспрессии рекомбинантных генов в клетке-реципиенте нужно использовать поксвирусные промоторы [42]. Поксвирусные транскрипты не подвергаются сплайсингу, из-за чего ДНК интереса обязательно должна быть представлена в форме комплементарной ДНК. Особенностью жизненного цикла поксвирусов является наличие собственной ДНК-зависимой РНК-полимеразы, которая обеспечивает считывание более половины вирусного генома в течение начальной и ранней стадий репродуктивного цикла. Вследствие большого размера и неинфекционной природы поксвирусной ДНК чужеродные гены клонируются в поксвирусах путем рекомбинации в условиях in vivo. Поксвирусы обладают природным тропизмом к опухолевой ткани [43].
Главными недостатками поксвирусов являются высокая иммуногенность, кратковременная продолжительность экспрессии и сложности репликации в эукариотических клетках. При этом положительными сторонами поксвирусов служат высокая эффективность трансдукции, емкость и способность заражать большинство типов клеток [44].
Аденоассоциированные вирусные системы и их преимущества
Рекомбинантные аденоассоциированные вирусы (ААВ) являются одними из наиболее перспективных векторов доставки для генной терапии и нейробиологии благодаря своим непатогенным свойствам, низкой иммуногенности со стороны хозяина, тропности к большинству клеток и тканей, высокой эффективности трансдукции и продолжительной экспрессии [45]. Основной их недостаток — небольшая емкость вектора (до 5 тыс. пар оснований).
Всего у ААВ известно 8 различных серотипов. ААВ2, ААВ3 и ААВ5 первоначально были выделены у обезьян [46]. Полагают, что ААВ6 был сформирован путем рекомбинирования ААВ2 и ААВ1, а именно 5’-конец ААВ2 слился с 3’-концом ААВ1 (включая две открытые рамки считывания) [47, 48]. ААВ7 и ААВ8 были выделены от макаки-резуса при ПЦР-амплификации с использованием праймеров, полученных из консервативных областей серотипов ААВ1–ААВ6 [49]. Среди всех восьми серотипов ААВ2 является наиболее изученным и широко используется в качестве вектора доставки генов [5].
ААВ принадлежит к семейству Parvoviridae. Это группа безоболочечных вирусов, содержащих одноцепочечную ДНК. Вирусная частица имеет икосаэдрическую (20 граней) симметрию с диаметром 18–26 нм и молекулярную массу 5,5–6,2 МДа [50].
Наиболее характерными особенностями ядерной структуры являются группы тройных выступов и выступающих петель, расположенных между двумя смежными субъединицами [51]. Положительно заряженные группы, расположенные вдоль одной стороны каждого выступа, отвечают за связывание с клеточным рецептором — гепарансульфат протеогликаном (HSPG) [52]. В связи с высокой экспрессией данного рецептора в разных тканях связывание с HSPG определяет широкий спектр клеточной специфичности к ААВ2-инфекции [53]. Петля, расположенная в соседней субъединице, также участвует в связывании с клеточным рецептором HSPG и содержит эпитоп для нейтрализующих антител [5]. Нейтрализация иммунного ответа определяет возможность будущего успешного повторного попадания вирусных векторов. Анализ структуры вирусных частиц ААВ2 с помощью рентгеновского излучения открыл возможность изменения эпитопа, содержащегося в вирусных белках капсида, и нейтрализующего антитела и осуществления других модификаций, применимых для реадмиссии в генной терапии [54].
ААВ2 имеет одноцепочечную геномную ДНК, состоящую из 4680 оснований. Инвертированные концевые повторы, состоящие из 145 оснований с высоким содержанием пар GC и способные образовывать палиндромную структуру, расположены как на 5’-конце, так и на 3’-конце [55]. ААВ2-геном синтезируется с мРНК, начиная от промотора p19 [56]. Ген cap кодирует три вирусных белка: VP1 (735 аминокислот, 90 кДа), VP2 (598 аминокислот, 72 кДа) и VP3 (533 аминокислот, 60 кДа), образующих вирусный капсид в соотношении 1:1:20 соответственно, при этом капсид имеет икосаэдрическую симметрию и состоит из 60 субъединиц [57]. Три капсидных белка синтезируются с одной мРНК, начиная с промотора р40, и отличаются друг от друга благодаря альтернативному сплайсингу и разным кодонам инициации [58]. Все три белка капсида имеют одни и те же карбоксиконцевые последовательности и стоп-кодоны (рис. 1).
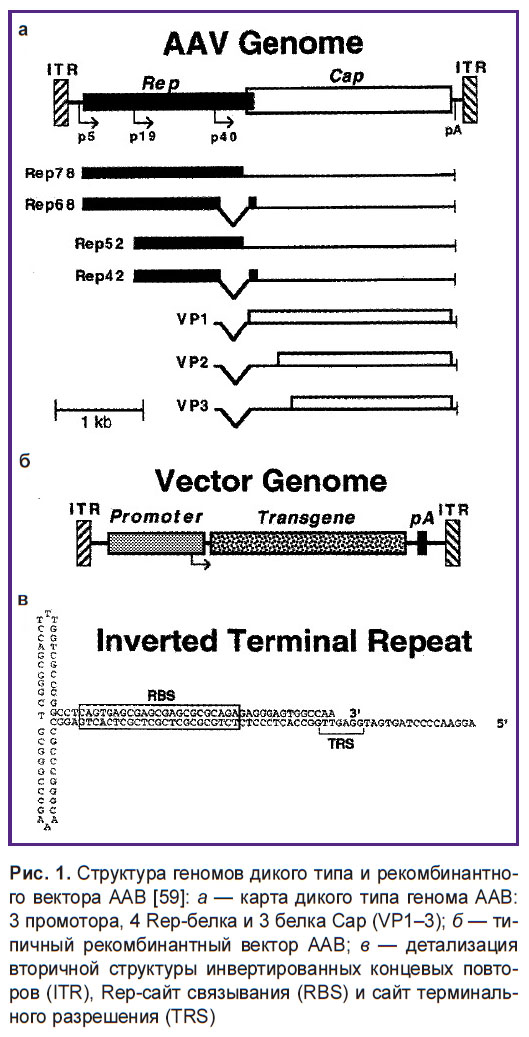
|
Рис. 1. Структура геномов дикого типа и рекомбинантного вектора ААВ [59]: а — карта дикого типа генома ААВ: 3 промотора, 4 Rep-белка и 3 белка Cap (VP1–3); б — типичный рекомбинантный вектор ААВ; в — детализация вторичной структуры инвертированных концевых повторов (ITR), Rep-сайт связывания (RBS) и сайт терминального разрешения (ТRS) |
Инвертированные концевые повторы на обоих концах выполняют по крайней мере следующие три функции. Во-первых, 3’-конец служит в качестве праймера для синтеза новой нити ДНК. Во-вторых, он имеет Rep-связывающий сайт (RBS) для Rep78 и Rep68, которые обладают хеликазной, цепьспецифической и сайтспецифической эндонуклеазной активностью [60]. В-третьих, там находится концевой сайт прикрепления (ТRS), который идентичен последовательности в 19-й хромосоме и выступает в качестве последовательности для интеграции с вирусным геномом [61].
Инфекционный путь ААВ2 включает несколько стадий (рис. 2). Вирусные частицы сначала прикрепляются к клеточной поверхности путем связывания с рецептором и корецептором, что в свою очередь ведет к интернализации и включению во внутриклеточный транспорт [62]. Затем эти частицы проникают через ядерную мембрану и либо интегрируют свои геномы с геномом клетки-хозяина, либо реплицируют свой геном внутри ядра [63]. Общепризнано, что основным клеточным рецептором для связывания ААВ2 является HSPG [64].
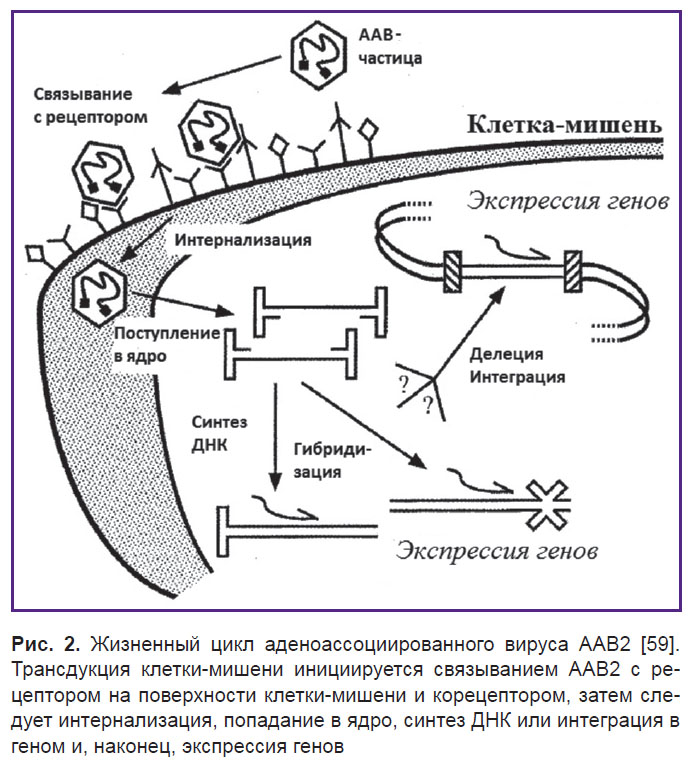
|
Рис. 2. Жизненный цикл аденоассоциированного вируса ААВ2 [59]. Трансдукция клетки-мишени инициируется связыванием ААВ2 с рецептором на поверхности клетки-мишени и корецептором, затем следует интернализация, попадание в ядро, синтез ДНК или интеграция в геном и, наконец, экспрессия генов |
Благодаря анализу сайтнаправленного мутагенеза была обнаружена важность основных аминокислот в связывании капсида ААВ2 с гепарином [65].
HSPG выступает в качестве основного рецептора, но ААВ2 также связывается с корецепторами — рецептором 1 фактора роста фибробластов (FGFR1) и αvβ5-интегрином [66, 67]. FGFR1, как полагают, увеличивает степень прикрепления вируса к клетке, в то время как αvβ5-интегрин необходим при эндоцитозе [68, 69]. После эндоцитоза вирусные частицы ААВ2 освобождаются от эндосомы при низком рН [70, 71]. Низкий рН, вероятно, вызывает конформационные изменения вирусных белков, которые играют решающую роль в успешном выходе из эндосомы и проникновении в ядро [72, 73].
В одних ядрах геном ААВ интегрирует в 19-ю хромосому в качестве провируса, а в других вирусные геномы могут оставаться как эписомы [5].
Хелперные плазмиды. Трансфекция
Трансфекция — процесс введения нуклеиновой кислоты в клетки человека и животных невирусным методом. Плазмидную ДНК наиболее часто трансфецируют в клетки, но и другие макромолекулы могут также быть введены внутрь клетки, такие как малые интерферирующие РНК, олигонуклеотиды, РНК и белки [74]. Трансфекция может состоять из нескольких стадий, но обязательным этапом является образование в плазматической мембране пор, через которые внутрь клетки способен проникать внеклеточный материал [75, 76].
Стандартный метод получения рААВ включает в себя котрансфекцию клеток рААВ плазмидой и упаковочной плазмидой, кодирующей гены Rep и белки VP (variol protein) [77]. Кроме того, клетки могут быть коинфицированы вирусом-помощником, например аденовирусом, который обеспечивает выполнение различных функций в производстве ААВ [78, 79]. Как следствие, клетки производят не только ААВ, но и аденовирусы, которые загрязняют конечный продукт (рис. 3, а).
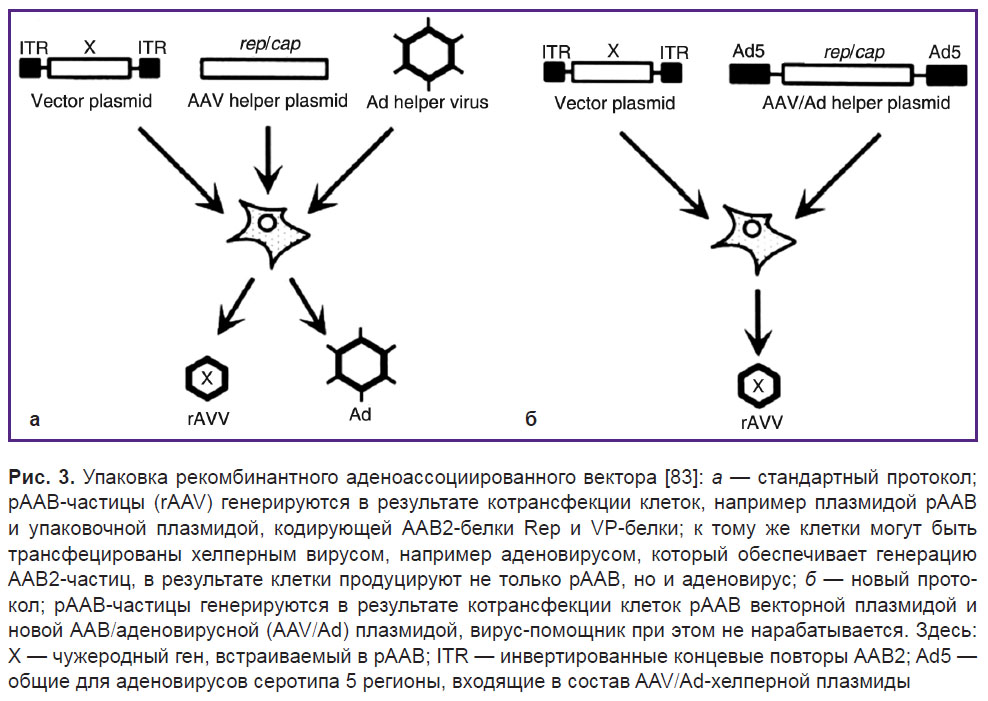
|
Рис. 3. Упаковка рекомбинантного аденоассоциированного вектора [83]: а — стандартный протокол; рААВ-частицы (rAAV) генерируются в результате котрансфекции клеток, например плазмидой рААВ и упаковочной плазмидой, кодирующей ААВ2-белки Rep и VP-белки; к тому же клетки могут быть трансфецированы хелперным вирусом, например аденовирусом, который обеспечивает генерацию ААВ2-частиц, в результате клетки продуцируют не только рААВ, но и аденовирус; б — новый протокол; рААВ-частицы генерируются в результате котрансфекции клеток рААВ векторной плазмидой и новой ААВ/аденовирусной (AAV/Ad) плазмидой, вирус-помощник при этом не нарабатывается. Здесь: Х — чужеродный ген, встраиваемый в рААВ; ITR — инвертированные концевые повторы ААВ2; Ad5 — общие для аденовирусов серотипа 5 регионы, входящие в состав AAV/Ad-хелперной плазмиды |
Передовой же метод получения рААВ основан на котрансфекции клеток рААВ и хелперной плазмидой, полученной из ААВ и аденовируса [80–82]. При этом производства аденовируса не происходит (рис. 3, б) [83].
Хелперная плазмида, или плазмида-помощник, обладает одной или несколькими функциями, которые отсутствуют у дефектной плазмиды [84]. Трансформированные клетки должны быть инфицированы также вспомогательным вирусом, например аденовирусом, что обеспечивает множество необходимых преимуществ для производства рААВ [85–87].
Типичные методы производства рААВ-векторов требуют котрансфекции клеток плазмидным вектором ААВ2 и вспомогательной плазмидой (хелперной) [88].
Хелперная плазмида экспрессирует гены rep и cap, а также гены, кодирующие продукты, важные для производства ААВ2 (рис. 4). Простой котрансфекции клеток с хелперной плазмидой и ААВ2-вектором достаточно для успешной наработки рААВ [83, 89].
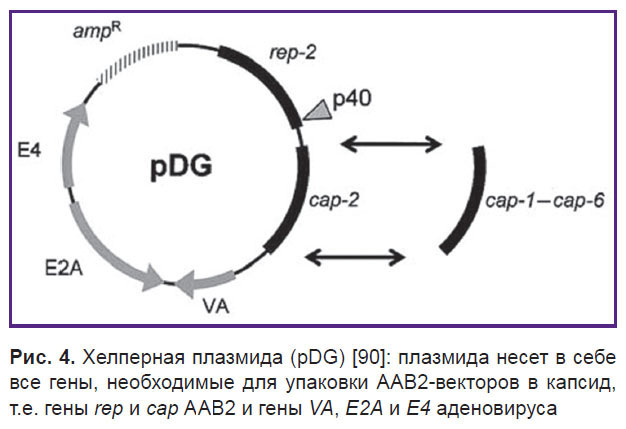 |
Рис. 4. Хелперная плазмида (pDG) [90]: плазмида несет в себе все гены, необходимые для упаковки ААВ2-векторов в капсид, т.е. гены rep и cap АAВ2 и гены VA, E2A и E4 аденовируса |
Рекомбинантные векторы, полученные из ААВ2, — перспективный инструмент для генной терапии и нейробиологии, так как они являются непатогенными и могут долгосрочно экспрессироваться в различных тканях [91–93].
Основные свойства отдельных вирусных векторов, такие как размер вирусных частиц, размер генома и емкость вставки, иммуногенность, а также эффективность трансдукции и экспрессии вируса в клетках-мишенях, отражены в таблице (см. таблицу).
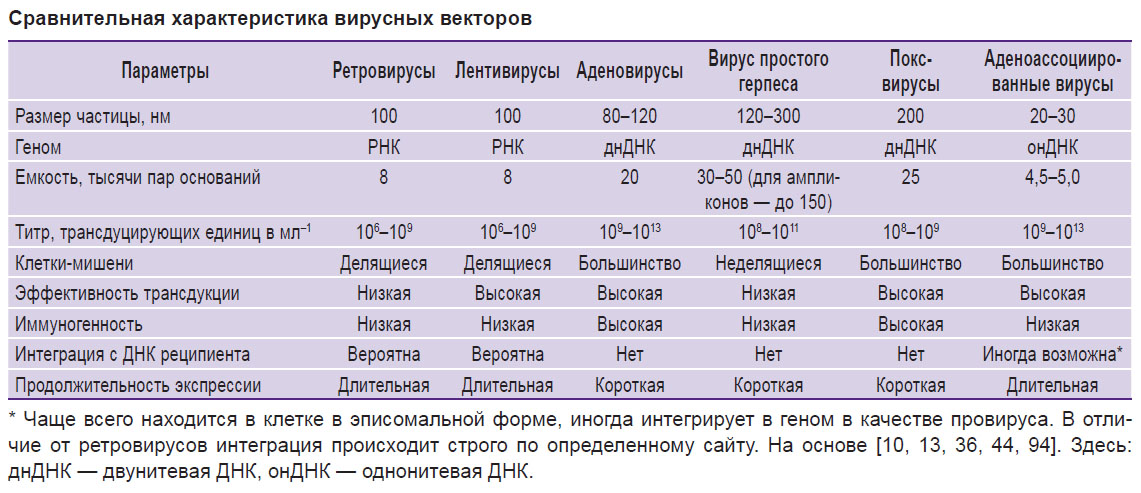 Сравнительная характеристика вирусных векторов Сравнительная характеристика вирусных векторов
|
Применение генетических вирусных векторов в нейробиологии
Перенос генетической информации в клетки центральной нервной системы имеет большие перспективы для фундаментальных и клинических исследований в нейробиологии [95, 96]. Так как мозг представляет собой сложноорганизованную структуру, это делает затруднительным генетические манипуляции в нем [97]. В настоящее время вирусные векторы являются наиболее широко используемыми транспортными средствами для переноса генов в зрелые клетки мозга млекопитающих [98, 99]. Тем не менее не существует «универсального идеального вектора» и любые исследования мозга требуют использования определенных векторных систем [100–102]. Кроме того, все вирусные векторные системы имеют свои недостатки и преимущества в зависимости от выбранных клеток-мишеней и специфики каждого исследования [103]. Причем количество векторов, подходящих для фундаментальных исследований, превосходит количество тех, которые могут применяться в клинических испытаниях [104, 105]. Сейчас наиболее широко используемыми векторами в нейробиологии являются аденоассоциированные, аденовирусные, герпесвирусные и лентивирусные векторы [106].
Рекомбинантные ААВ представляют собой векторы нескольких серотипов со специфическим клеточным тропизмом. Например, AАВ2 заражает преимущественно нейроны, но не все виды нейронов заражаются одинаково хорошо. Другие серотипы, такие как ААВ4 и ААВ5, демонстрируют различный тропизм и способность к диффузии [25].
Cre-рекомбиназа в сочетании с Cre-зависимой экспрессией белков и рААВ используется для достижения экспрессии трансгена в различных нейронных популяциях. При этом Cre-зависимые векторы показали надежную способность к ретроградной инфекции и экспрессии трансгена [82].
Генетически кодируемые нейрональные сайленсеры используются для проверки участия нейрональных популяций в поведенческих функциях [107]. Например, соматостатин-экспрессирующие клетки (SST) в комплексе Бетцингера (структура заднего мозга, которая регулирует дыхание) селективно выключались, чтобы проверить их участие в дыхании [108]. В этом исследовании использовались G-белки, связывающиеся с рецепторами аллатостатина (AlstR) дрозофилы, которые могут заставить «замолчать» нейроны млекопитающих при образовании комплекса рецептор–лиганд путем активации проводимости калия. Отдельно лиганд и его рецептор, как полагают, не могут повлиять на нейроны млекопитающих. Использование рААВ2 позволило добиться специфической экспрессии AlstR в субпопуляции клеток SST.
Оценка электрической активности нейронов широко исследовалась для изучения функции нейронов, а в последнее время — для характеристики нейронных сетей. Мониторинг активности нейрона — сложная задача, которая чаще всего сводится к крупномасштабной, мультинейронной визуализации уровня кальция, отражающего электрическую активность [109]. Рекомбинантные ААВ могут доставить генетически закодированные индикаторы кальция для визуализации динамики кальция в естественных условиях с достаточными уровнями экспрессии для эффективного обнаружения. Так, рААВ-экпрессия кальциевого индикатора camgaroo-2 под контролем тетрациклинового трансактиваторного промоторного элемента была осуществлена в обонятельной луковице трансгенных мышей CaMKIIa-tTA [110]. Это вызвало экспрессию в обонятельных сенсорных аксонах и позволило контролировать динамику кальция в ответ на различные ароматические вещества.
Другой кальциевый индикатор GCaMP3 был экспрессирован в клетках моторной коры мыши с использованием рAAВ2/рААВ5, что позволило контролировать динамическую активность нескольких нейронов у бегущей мыши [111].
Также ААВ применяются для выявления и оценки синаптических связей между нейронами разных типов. К примеру, такой подход был использован, чтобы отличить проекционные модели двух смешанных нейронных популяций, которые различаются по экспрессии типов дофаминовых рецепторов 1 или 2 (D1R или D2R) [109]. Применение Cre-зависимого вирусного вектора и сильного промотора EF-1α позволило управлять GFP-трансгеном в сочетании с наличием генетической модификации в мышах, что раскрыло поразительно четкие различия стрионигральной и стриопаллидарной субстанций [111].
Ампликоны вируса простого герпеса применяются для экспрессии генов большого размера или генов с повышенной копийностью. Так, например, мутантная форма торсина А, который ответственен за развитие торсионной дистрофии и формирует витые включения в цитоплазматической мембране [112], может быть экспрессирована в культуре клеток глиомы с помощью ампликона вируса простого герпеса [113].
В лабораторной практике применяются лентивирусные векторы с синапсиновым промотором, способствующим экспрессии заданного гена в любых типах нейрональных клеток [114–117]. Также используются лентивирусные векторы с промотором, содержащим кальций-кальмодулин-зависимую протеинкиназу II альфа, с экспрессией в возбуждающих глутаматергических нейронах [118, 119]. Применяются данные векторы для оптического и электрофизиологического мониторинга их активности, а также для оптогенетической стимуляции, например при остром паркинсонизме, индуцированном посредством медикаментов.
Аденовирусные векторы также применяются в нейробиологии, например в оптогенетике. Так, лабораторией S. Kasparova был создан оптогенетический активатор, который управляет астроцитарной активностью через экспрессию разных вариантов чанелродопсина-2 (ChR2), с целью запуска внеклеточных событий, таких как внеклеточное увеличение концентрации ионов Ca2+ [120].
Заключение
На данный момент не существует универсального «идеального» вектора, и разные исследования требуют использования определенных векторных систем. У всех вирусных векторных систем будут свои недостатки и преимущества в зависимости от выбираемых клеток-мишеней и специфики каждого исследования. В частности, преимущества рекомбинантных аденоассоциированных векторов (способность интегрировать целевой ген в геном хозяина в нужное место, что предотвращает нежелательные мутации; встраивание как в делящиеся, так и в покоящиеся клетки; широкий профиль трансдукции; низкий иммунный ответ; сильная и устойчивая экспрессия трансгена) выделяют их среди остальных вирусных векторов и делают эти векторы популярным и универсальным инструментом для доставки гена in vitro и in vivo.
Финансирование исследования. Работа выполнена при поддержке Федеральной целевой программы «Исследования и разработки по приоритетным направлениям развития научно-технологического комплекса России на 2014–2020 годы» Министерства образования и науки России, соглашение 14.581.21.0016 («Разработка методов, технологий и платформ для исследований функционирования нервных систем на основе создания высокоразрешающей информационной модели кортикальных структур мозга», уникальный идентификатор ПНИЭР RFMEFI58115X0016).
Конфликт интересов. Авторы декларируют отсутствие конфликта интересов.
Литература
- Daya S., Berns K.I. Gene therapy using adeno-associated virus vectors. Clin Microbiol Rev 2008; 21(4): 583–593, https://doi.org/10.1128/cmr.00008-08.
- During M.J., Leone P. Adeno-associated virus vectors for gene therapy of neurodegenerative disorders. Clin Neurosci 1995; 3(5): 292–300.
- Goff S.P., Berg P. Construction of hybrid viruses containing SV40 and lambda phage DNA segments and their propagation in cultured monkey cells. Cell 1976; 9(4): 695–705, https://doi.org/10.1016/0092-8674(76)90133-1.
- Lu Y. Recombinant adeno-associated virus as delivery vector for gene therapy — a review. Stem Cells Dev 2004; 13(1): 133–145, https://doi.org/10.1089/154732804773099335.
- Seow Y., Wood M.J. Biological gene delivery vehicles: beyond viral vector. Mol Ther 2009; 17(5): 767–777, https://doi.org/10.1038/mt.2009.41.
- Новикова Н.А. Хранение и реализация генетической информации вирусов. Учебно-методический материал по программе повышения квалификации «Хранение и обработка информации в биологических системах». Н. Новгород; 2007.
- Nienhuis A.W. Development of gene therapy for blood disorders. Blood 2008; 111(9): 4431–4444, https://doi.org/10.1182/blood-2007-11-078121.
- McClure C., Cole K.L.H., Wulff P., Klugmann M., Murray A.J. Production and titering of recombinant adeno-associated viral vectors. J Vis Exp 2011; 57: e3348, https://doi.org/10.3791/3348.
- Davidson B.L., Stein C.S., Heth J.A., Martins I., Kotin R.M., Derksen T.A., Zabner J., Ghodsi A., Chiorini J.A. Recombinant adeno-associated virus type 2, 4, and 5 vectors: transduction of variant cell types and regions in the mammalian central nervous system. Proc Natl Acad Sci USA 2000; 97(7): 3428–3432, https://doi.org/10.1073/pnas.97.7.3428.
- Boeckle S., Wagner E. Optimizing targeted gene delivery: chemical modification of viral vectors and synthesis of artificial virus vector systems. AAPS J 2006; 8(4): E731–E742, https://doi.org/10.1208/aapsj080483.
- Giacca M., Zacchigna S. Virus-mediated gene delivery for human gene therapy. J Control Release 2012; 161: 377–388, https://doi.org/10.1016/j.jconrel.2012.04.008.
- Daniel R., Smith J.A. Integration site selection by retroviral vectors: molecular mechanism and clinical consequences. Hum Gen Ther 2007; 19: 557–556, https://doi.org/10.1089/hum.2007.148.
- Nemunaitis J., Edelman J. Selectively replicating viral vectors. Cancer Gene Therapy 2002; 9: 987–1000, https://doi.org/10.1038/sj.cgt.7700547.
- Verma I.M., Naldini L., Kafri T., Miyoshi H., Takahashi M., Blömer U., Somia N., Wang L., Gage F.H. Gene therapy: promises, problems and prospects. Nature 1997; 389(6648): 239–242, https://doi.org/10.1007/978-3-642-56947-0_13.
- Lode H.N., Reisfeld R.A. Targeted cytokines for cancer immunotherapy. Immunol Res 2000; 21(2–3): 279–288, https://doi.org/10.1385/ir:21:2-3:279.
- Jolly D. Viral vector systems for gene therapy. Cancer Gene Ther 1994; 1(1): 51–64.
- Verhoeyen E., Cosset F.L. Surface-engineering of lentiviral vectors. J Gene Med 2004; 6(S1): 83–94, https://doi.org/10.1002/jgm.494.
- Zhang X.Y., La Russa V.F., Reiser J. Transduction of bonemarrow-derived mesenchymal stem cells by using lentivirus vectors pseudotyped with modified RD114 envelope glycoproteins. J Virol 2004; 78(3): 1219–1229, https://doi.org/10.1128/jvi.78.3.1219-1229.2004.
- Federici T., Kutner R., Zhang X.Y., Kuroda H., Tordo N., Boulis N.M., Reiser J. Comparative analysis of HIV-1-based lentiviral vectors bearing lyssavirus glycoproteins for neuronal gene transfer. Genet Vaccines Ther 2009; 7(1), https://doi.org/10.1186/1479-0556-7-1.
- Tang Y.Q., Yuan J., Osapay G., Osapay K., Tran D., Miller C.J., Ouellette A.J., Selsted M.E. A cyclic antimicrobial peptide produced in primate leukocytes by the ligation of two truncated α-defensins. Science 1999; 286(5439): 498–502, https://doi.org/10.1126/science.286.5439.498.
- Frecha C., Szécsi J., Cosset F.L., Verhoeyen E. Strategies for targeting lentiviral vectors. Curr Gene Ther 2008; 8(6): 449–460, https://doi.org/10.2174/156652308786848003.
- Cockrell A.S., Kafri T. Gene delivery by lentivirus vectors. Mol Biotechnol 2007; 36(3): 184–204, https://doi.org/10.1007/s12033-007-0010-8.
- Pluta K., Kacprzak M.M. Use of HIV as a gene transfer vector. Acta Biochim Pol 2009; 56(4): 531–595.
- Cronin J., Zhang X.Y., Reiser J. Altering the tropism of lentiviral vectors through pseudotyping. Curr Gene Ther 2005; 5(4): 387–398, https://doi.org/10.2174/1566523054546224.
- Wanisch K., Yáñez-Muñoz R.J. Integration-deficient lentiviral vectors: a slow coming of age. Mol Ther 2009; 17(8): 1316–1332, https://doi.org/10.1038/mt.2009.122.
- Coura R. Viral vectors in neurobiology: therapeutic and research applications. In: Molecular Virology. Moses P. Adoga (editor). InTech; 2012; p. 75–89, https://doi.org/10.5772/32492.
- Russell W.C. Adenoviruses: update on structure and function. J Gen Virol 2009; 90(1): 1–20, https://doi.org/10.1099/vir.0.003087-0.
- Roberts D.M., Nanda A., Havenga M.J., Abbink P., Lynch D.M., Ewald B.A., Liu J., Thorner A.R., Swanson P.E., Gorgone D.A., Lifton M.A., Lemckert A.A., Holterman L., Chen B., Dilraj A., Carville A., Mansfield K.G., Goudsmit J., Barouch D.H. Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature 2006; 441(7090): 239–243, https://doi.org/10.1038/nature04721.
- Segerman A., Arnberg N., Erikson A., Lindman K., Wadell G. There are two different species B adenovirus receptors: sBAR, common to species B1 and B2 adenoviruses, and sB2AR, exclusively used by species B2 adenoviruses. J Virol 2003; 77(2): 1157–1162, https://doi.org/10.1128/jvi.77.2.1157-1162.2003.
- Pichla-Gollon S.L., Lin S.W., Hensley S.E., Lasaro M.O., Herkenhoff-Haut L., Drinker M., Tatsis N., Gao G.P., Wilson J.M., Ertl H.C., Bergelson J.M. Effect of preexisting immunity on an adenovirus vaccine vector: in vitro neutralization assays fail to predict inhibition by antiviral antibody in vivo. J Virol 2009; 83(11): 5567–5573, https://doi.org/10.1128/jvi.00405-09.
- Zaiss A.K., Machado H., Herschman H.R. The influence of innate and pre-existing immunity on adenovirus therapy. J Cell Biochem 2009; 108(4): 778–790, https://doi.org/10.1002/jcb.22328.
- Sakurai F. Development and evaluation of a novel gene delivery vehicle composed of adenovirus serotype 35. Biol Pharm Bull 2008; 31(10): 1819–1825, https://doi.org/10.1248/bpb.31.1819.
- Sharma A., Li X., Bangari D.S., Mittal S.K. Adenovirus receptors and their implications in gene delivery. Virus Res 2009; 143(2): 184–194, https://doi.org/10.1016/j.virusres.2009.02.010.
- Akli S., Caillaud C., Vigne E., Stratford-Perricaudet L.D., Poenaru L., Perricaudet M., Kahn A., Peschanski M.R. Transfer of a foreign gene into the brain using adenovirus vectors. Nat Genet 1993; 3(3): 224–228, https://doi.org/10.1038/ng0393-224.
- Davidson B.L., Allen E.D., Kozarsky K.F., Wilson J.M., Roessler B.J. A model system for in vivo gene transfer into the central nervous system using an adenoviral vector. Nat Genet 1993; 3(3): 219–223, https://doi.org/10.1038/ng0393-219.
- Hukkanen V. Herpesvirus vectors in gene therapy. Open Virol J 2010; 4(3): 94–95, https://doi.org/10.2174/1874357901004030094.
- Bienkowska-Szewczyk K., Szewczyk B. Expression of genes coding for viral glycoproteins in heterologous systems. Acta Biochim Pol 1999; 46(2): 325–339.
- Burton E.A., Fink D.J., Glorioso J.C. Gene delivery using herpes simplex virus vectors. DNA Cell Biol 2002; 21(12): 915–936, https://doi.org/10.1089/104454902762053864.
- Fink D.J., Glorioso J.C. Herpes simplex viral vectors in gene therapy. In: Encyclopedia of life sciences. Wiley-Blackwell; 2007, https://doi.org/10.1002/9780470015902.a0005739.pub2.
- Sena-Esteves M., Saeki Y., Fraefel C., Breakefield X.O. HSV-1 amplicon vectors — simplicity and versatility. Mol Ther 2000; 2(1): 9–15, https://doi.org/10.1006/mthe.2000.0096.
- Laquerre S., Argnani R., Anderson D.B., Zucchini S., Manservigi R., Glorioso J.C. Heparan sulfate proteoglycan binding by herpes simplex virus type 1 glycoproteins B and C, which differ in their contributions to virus attachment, penetration, and cell-to-cell spread. J Virol 1998; 72(7): 6119–6130.
- Sandvik T., Tryland M., Hansen H., Mehl R., Moens U., Olsvik O., Traavik T. Naturally occurring orthopoxviruses: potential for recombination with vaccine vectors. J Clin Microbiol 1998; 36(9): 2542–2547.
- Zeh H.J., Bartlett D.L. Development of a replication-selective, oncolytic poxvirus for the treatment of human cancers. Cancer Gene Ther 2002; 9(12): 1001–1012, https://doi.org/10.1038/sj.cgt.7700549.
- Davidson B.L., Breakefield X.O. Viral vectors for gene delivery to the nervous system. Nat Rev Neurosci 2003; 4(5): 353–364, https://doi.org/10.1038/nrn1104.
- Somia N., Verma I.M. Gene therapy: trials and tribulations. Nat Rev Genet 2000; 1(2): 91–99, https://doi.org/10.1038/35038533.
- Berns K.I. Parvoviridae. The viruses and their replication. In: Virology. Fields B.N., Knipe D.M., Howley P.M. (editors). 3rd ed. Lippincott Williams and Wilkins, Philadelphia; 1996; p. 2173–2197.
- Rutledge E.A., Halbert C.L., Russell D.W. Infectious clones and vectors derived from adeno-associated virus (AAV) serotypes other than AAV type 2. J Virol 1998; 72(1): 309–319.
- Xiao W., Chirmule N., Berta S.C., McCullough B., Gao G., Wilson J.M. Gene therapy vectors based on adenoassociated virus type 1. J Virol 1999; 73(5): 3994–4003.
- Gao G.P., Alvira M.R., Wang L., Calcedo R., Johnston J., Wilson J.M. Novel adeno-associated viruses from rhesus monkeys as vectors for human gene therapy. Proc Natl Acad Sci USA 2002; 99(18): 11854–11859, https://doi.org/10.1073/pnas.182412299.
- Zhou X., Muzyczka N. In vitro packaging of adeno-associated virus DNA. J Virol 1998; 72: 3241–3247.
- Zolotukhin S., Byrne B.J., Mason E., Zolotukhin I., Potter M., Chesnut K., Summerford C., Samulski R.J., Muzyczka N. Recombinant adeno-associated virus purification using novel methods improves infectious titer and yield. Gene Ther 1999; 6(6): 973–985, https://doi.org/10.1038/sj.gt.3300938.
- Clark K.R., Liu X., McGrath J.P., Johnson P.R. Highly purified recombinant adeno-associated virus vectors are biologically active and free of detectable helper and wild-type viruses. Hum Gene Ther 1999; 10(6): 1031–1039, https://doi.org/10.1089/10430349950018427.
- Young S.M., Samulski R.J. Adeno-associated virus (AAV) site-specific recombination does not require a Rep-dependent origin of replication within the AAV terminal repeat. Proc Natl Acad Sci USA 2001; 98(24): 13525–13530, https://doi.org/10.1073/pnas.241508998.
- Douar A.M., Poulard K., Stockholm D., Danos O. Intracellular trafficking of adeno-associated virus vectors: routing to the late endosomal compartment and proteasome degradation. J Virol 2001; 75(4): 1824–1833, https://doi.org/10.1128/jvi.75.4.1824-1833.2001.
- Sun J.Y., Anand-Jawa V., Chatterjee S., Wong K.K. Immune responses to adeno-associated virus and its recombinant vectors. Gene Ther 2003; 10(11): 964–976, https://doi.org/10.1038/sj.gt.3302039.
- Grimm D. Production methods for gene transfer vectors based on adeno-associated virus serotypes. Methods 2002; 28(2): 146–157, https://doi.org/10.1016/s1046-2023(02)00219-0.
- Büning H., Ried M.U., Perabo L., Gerner F.M., Huttner N.A., Enssle J., Hallek M. Receptor targeting of adeno-associated virus vectors. Gene Ther 2003; 10(14): 1142–1151, https://doi.org/10.1038/sj.gt.3301976.
- Balagúe C., Kalla M., Zhang W.W. Adeno-associated virus Rep78 protein and terminal repeats enhance integration of DNA sequences into the cellular genome. J Virol 1997; 71(4): 3299–3306.
- Russell D.W., Kay M.A. Adeno-associated virus vectors and hemalology. Blood 1999; 94(3): 864–874.
- Tsai T.H., Chen S.L., Xiao X., Liu D.W., Tsao Y.P. Gene therapy for treatment of cerebral ischemia using defective recombinant adeno-associated virus vectors. Methods 2002; 28(2): 253–258, https://doi.org/10.1016/s1046-2023(02)00230-x.
- Young S.M., McCarty D.M., Degtyareva N., Samulski R.J. Roles of adeno-associated virus Rep protein and human chromosome 19 in site-specific recombination. J Virol 2000; 74(9): 3953–3966, https://doi.org/10.1128/jvi.74.9.3953-3966.2000.
- Hildinger M., Auricchio A., Gao G., Wang L., Chirmule N., Wilson J.M. Hybrid vectors based on adeno-associated virus serotypes 2 and 5 for muscle-directed gene transfer. J Virol 2001; 75(13): 6199–6203, https://doi.org/10.1128/jvi.75.13.6199-6203.2001.
- Chao H., Liu Y., Rabinowitz J., Li C., Samulski R.J., Walsh C.E. Several log increase in therapeutic transgene delivery by distinct adeno-associated viral serotype vectors. Mol Ther 2000; 2(6): 619–623, https://doi.org/10.1006/mthe.2000.0219.
- Snyder R.O. Adeno-associated virus-mediated gene delivery. J Gene Med 1999; 1(3): 166–175, https://doi.org/10.1002/(sici)1521-2254(199905/06...>3.0.co;2-z.
- Wu P., Xiao W., Conlon T., Hughes J., Agbandje-McKenna M., Ferkol T., Flotte T., Muzyczka N. Mutational analysis of the adeno-associated virus type 2 (AAV2) capsid gene and construction of AAV2 vectors with altered tropism. J Virol 2000; 74(18): 8635–8647, https://doi.org/10.1128/jvi.74.18.8635-8647.2000.
- Xie Q., Bu W., Bhatia S., Hare J., Somasundaram T., Azzi A., Chapman M.S. The atomic structure of adeno-associated virus (AAV-2), a vector for human gene therapy. Proc Natl Acad Sci USA 2002; 99(16): 10405–10410, https://doi.org/10.1073/pnas.162250899.
- Xie Q., Somasundaram T., Bhatia S., Bu W., Chapman M.S. Structure determination of adeno-associated virus 2: three complete virus particles per asymmetric unit. Acta Crystallogr D Biol Crystallogr 2003; 59(6): 959–970, https://doi.org/10.1107/s0907444903005675.
- Doria M., Ferrara A., Auricchio A. AAV2/8 vectors purified from culture medium with a simple and rapid protocol transduce murine liver, muscle, and retina efficiently. Hum Gene Ther Method 2013; 24(6): 392–398, https://doi.org/10.1089/hgtb.2013.155.
- Mietzsch M., Grasse S., Zurawski C., Weger S., Bennett A., Agbandje-McKenna M., Muzyczka N., Zolotukhin S., Heilbronn R. OneBac: platform for scalable and high-titer production of adeno-associated virus serotype 1–12 vectors for gene therapy. Hum Gene Ther 2014; 25(3): 212–222, https://doi.org/10.1089/hum.2013.184.
- Grieger J.C., Samulski R.J. Adeno-associated virus vectorology, manufacturing, and clinical applications. Methods Enzymol 2012; 507: 229–254, https://doi.org/10.1016/b978-0-12-386509-0.00012-0.
- Jooss K., Chirmule N. Immunity to adenovirus and adeno-associated viral vectors: implications for gene therapy. Gene Ther 2003; 10(11): 955–963, https://doi.org/10.1038/sj.gt.3302037.
- Nair H.P., Young L.J. Application of adeno-associated viral vectors in behavioral research. Methods 2002; 28(2): 195–202, https://doi.org/10.1016/s1046-2023(02)00223-2.
- Ponnazhagan S., Mahendra G., Kumar S., Thompson J.A., Castillas M. Conjugate-based targeting of recombinant adeno-associated virus type 2 vectors by using avidin-linked ligands. J Virol 2002; 76(24): 12900–12907, https://doi.org/10.1128/jvi.76.24.12900-12907.2002.
- Peel A.L., Zolotukhin S., Schrimsher G.W., Muzyczka N., Reier P.J. Efficient transduction of green fluorescent protein in spinal cord neurons using adeno-associated virus vectors containing cell type-specific promoters. Gene Ther 1997; 4(1): 16–24, https://doi.org/10.1038/sj.gt.3300358.
- Groskreutz D., Schenborn E.T. Reporter systems. In: Recombinant protein protocols. Methods in molecular biology. Springer Nature; 1997; p. 11–30, https://doi.org/10.1385/0-89603-481-x:11.
- El-Andaloussi N., Endele M., Leuchs B., Bonifati S., Kleinschmidt J., Rommelaere J., Marchini A. Novel adenovirus-based helper system to support production of recombinant parvovirus. Cancer Gene Ther 2010; 18(4): 240–249, https://doi.org/10.1038/cgt.2010.73.
- Rabinowitz J.E., Rolling F., Li C., Conrath H., Xiao W., Xiao X., Samulski R.J. Cross-packaging of a single adeno-associated virus (AAV) type 2 vector genome into multiple AAV serotypes enables transduction with broad specificity. J Virol 2002; 76(2): 791–801, https://doi.org/10.1128/jvi.76.2.791-801.2002.
- Dubielzig R., King J.A., Weger S., Kern A., Kleinschmidt J.A. Adeno-associated virus type 2 protein interactions: formation of pre-encapsidation complexes. J Virol 1999; 73: 8989–8998.
- Wistuba A., Kern A., Weger S., Grimm D., Kleinschmidt J.A. Subcellular compartmentalization of adeno-associated virus type 2 assembly. J Virol 1997; 71(2): 1341–1352.
- Holehonnur R., Luong J.A., Chaturvedi D., Ho A., Lella S.K., Hosek M.P., Ploski J.E. Adeno-associated viral serotypes produce differing titers and differentially transduce neurons within the rat basal and lateral amygdala. BMC Neurosci 2014; 15(1): 28, https://doi.org/10.1186/1471-2202-15-28.
- Nicolas A., Jolinon N., Alazard-Dany N., Barateau V., Epstein A.L., Greco A., Büning H., Salvetti A. Factors influencing helper-independent adeno-associated virus replication. Virology 2012; 432(1): 1–9, https://doi.org/10.1016/j.virol.2012.05.027.
- Rothermel M., Brunert D., Zabawa C., Díaz-Quesada M., Wachowiak M. Transgene expression in target-defined neuron populations mediated by retrograde infection with adeno-associated viral vectors. J Neurosci 2013; 33(38): 15195–15206, https://doi.org/10.1523/jneurosci.1618-13.2013.
- Grimm D., Kern A., Ritter K., Kleinschmedt J.A. Novel tools for production and purification of recombinant adenoassociated virus vectors. Hum Gene Ther 1998; 9(18): 2745–2760, https://doi.org/10.1089/hum.1998.9.18-2745.
- Kotin R.M. Large-scale recombinant adeno-associated virus production. Hum Mol Genet 2011; 20(R1): R2–R6, https://doi.org/10.1093/hmg/ddr141.
- Khan I.F., Hirata R.K., Russell D.W. AAV-mediated gene targeting methods for human cells. Nat Protoc 2011; 6(4): 482–501, https://doi.org/10.1038/nprot.2011.301.
- Dong B., Moore A.R., Dai J., Roberts S., Chu K., Kapranov P., Moss B., Xiao W. A concept of eliminating nonhomologous recombination for scalable and safe AAV vector generation for human gene therapy. Nucleic Acids Res 2013; 41(13): 6609–6617, https://doi.org/10.1093/nar/gkt404.
- Chiorini J.A., Afione S., Kotin R.M. Adeno-associated virus (AAV) type 5 Rep protein cleaves a unique terminal resolution site compared with other AAV serotypes. J Virol 1999; 73(5): 4293–4298.
- Kay M.A., Manno C.S., Ragni M.V., Larson P.J., Couto L.B., McClelland A., Glader B., Chew A.J., Tai S.J., Herzog R.W., Arruda V., Johnson F., Scallan C., Skarsgard E., Flake A.W., High K.A. Evidence for gene transfer and expression of factor IX inhaemophilia B patients treated with an AAV vector. Nat Genet 2000; 24(3): 257–261, https://doi.org/10.1038/73464.
- Grimm D. Adeno-associated virus (AAV) serotypes as vectors for human gene therapy. In: Research advances in virology. Vol. 1. Samal S.K., Liebler-Tenorio E., Delsert C., Paton D. (editors). India: Global Research Network; 2000; p. 91–114.
- Grimm D., Kay M.A., Kleinschmidt J.A. Helper virus-free, optically controllable, and two-plasmid-based production of adeno-associated virus vectors of serotypes 1 to 6. Mol Ther 2003; 7(6): 839–850, https://doi.org/10.1016/s1525-0016(03)00095-9.
- Synder R.O., Xiao X., Samulski R.J. Production of recombinant adeno-associated viral vectors. In: Current protocols in human genetics. Dracopoli N. (editor). New York: John Wiley; 1996; Chpt. 12.1; p. 1–20.
- Xu Z., Shi C., Qian Q. Scalable manufacturing methodologies for improving adeno-associated virus-based pharmaprojects. Chinese Science Bulletin 2014; 59(16): 1845–1855, https://doi.org/10.1007/s11434-014-0197-6.
- Wright J. Product-related impurities in clinical-grade recombinant AAV vectors: characterization and risk assessment. Biomedicines 2014; 2(1): 80–97, https://doi.org/10.3390/biomedicines2010080.
- Lentz T.B., Gray S.J., Samulski R.J. Viral vectors for gene delivery to the central nervous system. Neurobiol Dis 2012; 48(2): 179–188, https://doi.org/10.1016/j.nbd.2011.09.014.
- Kamimura K., Suda T., Zhang G., Liu D. Advances in gene delivery systems. Pharmaceut Med 2011; 25(5): 293–306, https://doi.org/10.1007/bf03256872.
- Auricchio A. Exchange of surface proteins impacts on viral vector cellular specificity and transduction characteristics: the retina as a model. Hum Mol Genet 2001; 10(26): 3075–3081, https://doi.org/10.1093/hmg/10.26.3075.
- Snyder R.O., Im D.S., Muzyczka N. Evidence for covalent attachment of the adeno-associated virus (AAV) rep protein to the ends of the AAV genome. J Virol 1990; 64(12): 6204–6213.
- Paterna J.-C., Büeler H. Recombinant adeno-associated virus vector design and gene expression in the mammalian brain. Methods 2002; 28(2): 208–218, https://doi.org/10.1016/s1046-2023(02)00225-6.
- Kaspar B.K. Retrograde viral delivery of igf-1 prolongs survival in a mouse ALS model. Science 2003; 301(5634): 839–842, https://doi.org/10.1126/science.1086137.
- Chatterjee S., Li W., Wong C.A., Fisher-Adams G., Lu D., Guha M., Macer J.A., Forman S.J., Wong K.K. Jr. Transduction of primitive human marrow and cord blood-derived hematopoietic progenitor cells with adeno-associated virus vectors. Blood 1999; 93: 1882–1894.
- Hermonat P.L., Muzyczka N. Use of adeno-associated virus as a mammalian DNA cloning vector: transduction of neomycin resistance into mammalian tissue culture cells. Proc Natl Acad Sci USA 1984; 81(20): 6466–6470, https://doi.org/10.1073/pnas.81.20.6466.
- Polinski N.K., Gombash S.E., Manfredsson F.P., Lipton J.W., Kemp C.J., Cole-Strauss A., Kanaan N.M., Steece-Collier K., Kuhn N.C., Wohlgenant S.L., Sortwell C.E. Recombinant adenoassociated virus 2/5-mediated gene transfer is reduced in the aged rat midbrain. Neurobiol Aging 2015; 36(2): 1110–1120, https://doi.org/10.1016/j.neurobiolaging.2014.07.047.
- Weger S., Wistuba A., Grimm D., Kleinschmidt J.A. Control of adeno-associated virus type 2 cap gene expression: relative influence of helper virus, terminal repeats, and Rep proteins. J Virol 1997; 71(11): 8437–8447.
- Tiesjema B., Adan R.A.H., Luijendijk M.C.M., Kalsbeek A., la Fleur S.E. Differential effects of recombinant adeno-associated virus-mediated neuropeptide Y overexpression in the hypothalamic paraventricular nucleus and lateral hypothalamus on feeding behavior. J Neurosci 2007; 27(51): 14139–14146, https://doi.org/10.1523/jneurosci.3280-07.2007.
- Nakai H., Storm T.A., Kay M.A. Recruitment of single-stranded recombinant adeno-associated virus vector genomes and intermolecular recombination are responsible for stable transduction of liver in vivo. J Virol 2000; 74(20): 9451–9463, https://doi.org/10.1128/jvi.74.20.9451-9463.2000.
- Kearns W.G., Afione S.A., Fulmer S.B., Pang M.C., Erikson D., Egan M., Landrum M.J., Flotte T.R., Cutting G.R. Recombinant adeno-associated virus (AAVCFTR) vectors do not integrate in a site-specific fashion in an immortahzed epithelial cell line. Gene Ther 1996; 3(9): 748–775.
- Betley J.N., Sternson S.M. Adeno-associated viral vectors for mapping, monitoring, and manipulating neural circuits. Hum Gene Ther 2011; 22(6): 669–677, https://doi.org/10.1089/hum.2010.204.
- Tan W., Janczewski W.A., Yang P., Shao X.M., Callaway E.M., Feldman J.L. Silencing preBötzinger Complex somatostatin-expressing neurons induces persistent apnea in awake rat. Nat Neurosci 2008; 11(5): 538–540, https://doi.org/10.1038/nn.2104.
- Tian L., Hires S.A., Mao T., Huber D., Chiappe M.E., Chalasani S.H., Petreanu L., Akerboom J., McKinney S.A., Schreiter E.R., Bargmann C.I., Jayaraman V., Svoboda K., Looger L.L. Imaging neural activity in worms, flies and mice with improved GCaMP calcium indicators. Nat Methods 2009; 6(12): 875–881, https://doi.org/10.1038/nmeth.1398.
- Hasan M.T., Friedrich R.W., Euler T., Larkum M.E., Giese G., Both M., Duebel J., Waters J., Bujard H., Griesbeck O., Tsien R.Y., Nagai T., Miyawaki A., Denk W. Functional fluorescent Ca2+ indicator proteins in transgenic mice under TET control. PLoS Biol 2004; 2(6): e163, https://doi.org/10.1371/journal.pbio.0020163.
- Kravitz A.V., Freeze B.S., Parker P.R., Kay K., Thwin M.T., Deisseroth K., Kreitzer A.C. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature 2010; 466(7306): 622–626, https://doi.org/10.1038/nature09159.
- Hewett J., Gonzalez-Agosti C., Slater D., Ziefer P., Li S., Bergeron D., Jacoby D.J., Ozelius L.J., Ramesh V., Breakefield X.O. Mutant torsinA, responsible for early-onset torsion dystonia, forms membrane inclusions in cultured neural cells. Hum Mol Genet 2000; 9(9): 1403–1413, https://doi.org/10.1093/hmg/9.9.1403.
- Bragg D.C., Wilbur J.D., Breakefield X.O. Expression of mutant and wild-type torsinA in human glioma cells by HSV amplicon vector-mediated gene transfer. In: Proc. 32nd Annual Meeting Society for Neuroscience. Orlando, Florida, USA; 2002.
- Hioki H., Kameda H., Nakamura H., Okunomiya T., Ohira K., Nakamura K., Kuroda M., Furuta T., Kaneko T. Efficient gene transduction of neurons by lentivirus with enhanced neuron-specific promoters. Gene Ther 2007; 14(11): 872–882, https://doi.org/10.1038/sj.gt.3302924.
- Nathanson J.L., Yanagawa Y., Obata K., Callaway E.M. Preferential labeling of inhibitory and excitatory cortical neurons by endogenous tropism of adeno-associated virus and lentivirus vectors. Neuroscience 2009; 161(2): 441–450, https://doi.org/10.1016/j.neuroscience.2009.03.032.
- Van Hooijdonk L.W., Ichwan M., Dijkmans T.F., Schouten T.G., de Backer M.W., Adan R.A., Verbeek F.J., Vreugdenhil E., Fitzsimons C.P. Lentivirus-mediated transgene delivery to the hippocampus reveals sub-field specific differences in expression. BMC Neurosci 2009; 10(1): 2, https://doi.org/10.1186/1471-2202-10-2.
- Yaguchi M., Ohashi Y., Tsubota T., Sato A., Koyano K.W., Wang N., Miyashita Y. Characterization of the properties of seven promoters in the motor cortex of rats and monkeys after lentiviral vector-mediated gene transfer. Hum Gene Ther Methods 2013; 24(6): 333–344, https://doi.org/10.1089/hgtb.2012.238.
- Dittgen T., Nimmerjahn A., Komai S., Licznerski P., Waters J., Margrie T.W., Helmchen F., Denk W., Brecht M., Osten P. Lentivirus-based genetic manipulations of cortical neurons and their optical and electrophysiological monitoring in vivo. Proc Natl Acad Sci USA 2004; 101(52): 18206–18211, https://doi.org/10.1073/pnas.0407976101.
- Seeger-Armbruster S., Bosch-Bouju C., Little S.T.C., Smither R.A., Hughes S.M., Hyland B.I., Parr-Brownlie L.C. Patterned, but not tonic, optogenetic stimulation in motor thalamus improves reaching in acute drug-induced parkinsonian rats. J Neurosci 2015; 35(3): 1211–1216, https://doi.org/10.1523/jneurosci.3277-14.2015.
- Figueiredo M., Lane S., Stout R.F., Liu B., Parpura V., Teschemacher A.G., Kasparov S. Comparative analysis of optogenetic actuators in cultured astrocytes. Cell Calcium 2014; 56(3): 208–214, https://doi.org/10.1016/j.ceca.2014.07.007.








