Влияние микро-РНК на активность сигнальных путей радиочувствительных и радиорезистентных раковых клеточных линий после радиационного облучения
Цель исследования — оценка экспрессии микро-РНК в радиорезистентных и радиочувствительных раковых клеточных линиях после радиационного облучения и изучение ее влияния на активность сигнальных путей.
Материалы и методы. В работе использовались радиорезистентная клеточная линия К562 и радиочувствительные клеточные линии HL-60 и Raji. Оценивали выживаемость клеток после облучения гамма-излучением 4 Гр. Изучали состав микро-РНК через 1, 4 и 24 ч после облучения c использованием метода параллельного массового секвенирования. Биоинформационную обработку полученных данных выполняли с использованием сервиса GenXPro и базы данных PANTHER.
Результаты. После однократного облучения клеток в дозе 4 Гр количество клеток с признаками некроза в радиочувствительных клеточных линиях увеличивается в несколько раз по сравнению с контролем. Микро-РНК hsa-miR-590-3p была найдена во всех исследуемых клеточных линиях на всех этапах эксперимента. Наиболее значительные отличия между радиорезистентной и радиочувствительными клеточными линиями наблюдаются в динамике влияния микро-РНК на сигнальные пути Integrin signaling pathway и General transcription by RNA polymerase I.
Заключение. Динамика и величина экспрессии микро-РНК hsa-miR-590-3p коррелируют с радиочувствительностью раковых клеток. Влияние ее на сигнальные пути радиорезистентных и радиочувствительных клеточных линий не одинаково: в радиорезистентной клеточной линии К562 ослаблено ингибирующее действие микро-РНК на Integrin signaling pathway и усилено — на General transcription by RNA polymerase I.
Радиорезистентность злокачественных опухолей является серьезной проблемой радиотерапии и служит основным препятствием для достижения максимальной эффективности лучевой терапии при лечении рака.
Радиорезистентность раковых клеток — это свойство, в основе которого лежит ряд генетических мутаций и нарушений нормальной экспрессии генов, что в свою очередь изменяет чувствительность раковых клеток к радиотерапии. Нарушения в экспрессии генов приводят к нарушениям в метаболизме клетки и изменениям регуляции клеточных процессов, в частности связанных с программируемой клеточной смертью. Феномен радиорезистентности формируется на надмолекулярном уровне и связан со способностью раковых клеток абортировать запуск механизмов программируемой клеточной смерти, индуцированных радиационным излучением [1, 2]. Механизмы программируемой клеточной смерти — апоптоз и автофагия — в нормальных клетках запускаются при невозможности репарации клеточных повреждений, в основном повреждений ДНК. Индукция программируемой клеточной смерти при стрессовых воздействиях не только связана со степенью повреждения ДНК, но и включает большое количество других сигнальных механизмов, а также зависит от их взаимодействия [3, 4]. Функционирование сигнальных и метаболических механизмов связано с регуляцией экспрессии генов на транскрипционном и посттранскрипционном уровнях.
Основным механизмом регуляции экспрессии генов на посттранскрипционном уровне является регуляция деградации матричной РНК (мРНК) посредством микро-РНК. Микро-РНК представляют собой небольшие (20–25 пар нуклеотидов) некодирующие РНК, влияющие на транскрипционную и посттранскрипционную регуляцию генов через подавление экспрессии [5, 6]. В геноме человека обнаружены более тысячи микро-РНК, каждая из которых потенциально может регулировать сотни мРНК, поэтому они играют определяющую роль во многих клеточных процессах, в том числе радиочувствительности и радиорезистентности клеток. Установлено [7], что микро-РНК регулируют экспрессию около 60% всех генов, кодирующих белок.
Исследования последних лет показали, что микро-РНК играют большую роль в патогенезе рака [8, 9]. Экспрессия их меняется под действием ионизирующего облучения, что может означать их участие в клеточном ответе на ионизирующее облучение [10]. Проведенные исследования [11–14] продемонстрировали связь некоторых микро-РНК с радиорезистентностью раковых клеток, что позволило выявить несколько десятков микро-РНК, дифференциально экспрессирующихся в радиорезистентных и радиочувствительных клеточных линиях. Однако не обнаружено таких микро-РНК, которые были бы характерны для всех радиорезистентных клеточных линий. Это связано с тем, что регуляция экспрессии генов посредством микро-РНК является сложным процессом, при котором одна микро-РНК может регулировать экспрессию многих генов, а экспрессию одного гена могут регулировать много разных микро-РНК. В этом случае более перспективным подходом в исследовании роли микро-РНК является изучение их влияния на сигнальные пути.
Цель исследования — оценка экспрессии микро-РНК в радиорезистентных и радиочувствительных раковых клеточных линиях после радиационного облучения и изучение ее влияния на активность сигнальных путей.
Материалы и методы. Эксперимент проводили на клеточных линиях HL-60 — промиелоцитарный лейкоз человека, К562 — радиорезистентная суспензионная линия хронической миелоидной лейкемии человека, Raji — линия лимфобластоидных клеток человека.
Клетки культивировали при температуре 37°С, в атмосфере 5% СО2 и при 98% влажности с использованием среды RPMI-1640, которая содержала L-глутамин с добавлением 10% эмбриональной бычьей сыворотки и 50 мкг/мл гентамицина.
Облучение клеточных линий проводили гамма-излучением во время логарифмической фазы роста на линейном ускорителе Elekta Synergy (Elekta, Швеция) в дозе 4 Гр (энергия фотонов 10 мэВ). Длительность облучения составляла 55 с при температуре 21–22°С. Поле облучения — 16×20 мм. Клетки находились в 6-луночных планшетах в количестве 5·105/мл среды, объем среды с клетками в каждой лунке составлял 2,5 мл.
Контрольные группы клеток не подвергались каким-либо воздействиям.
Выживаемость оценивали путем окраски клеток смесью флюоресцентных красителей акридинового оранжевого и пропидиум иодида. Окрашенные клетки анализировали на флюоресцентном микроскопе. Клетки с признаками некроза были окрашены в оранжевый цвет, живые клетки — в зеленый [15].
Общую РНК, обогащенную микро-РНК, выделяли из клеток с использованием набора Absolutely RNA miRNA Kit (Agilent Technologies, США) через 1, 4 и 24 ч после облучения. Качество выделенной РНК проверяли по соотношению 18S/28S РНК с использованием прибора для капиллярного электрофореза Agilent Bioanalyzer 2100 (Agilent Technologies, США). Для дальнейшей работы отбирали образцы с показателем целостности RIN>8,0. Получение библиотеки комплементарной ДНК (кДНК) микро-РНК, легирование с адаптерами и бар-кодирование проводили с использованием набора NEBNext Small RNA Library Prep Set (NEB, Великобритания). Очистку библиотек кДНК микро-РНК выполняли с помощью электрофореза с 6% полиакриламидным гелем. Фрагменты кДНК между 145 и 160 п. н., соответствующие микро-РНК, вырезали из геля, затем элюировали и осаждали 80% этанолом. Осадок кДНК высушивали на воздухе и ресуспендировали в 10 мкл воды, обработанной диэтилпирокарбонатом (DEPC). Количество кДНК в каждой из финальных библиотек микро-РНК оценивали с использованием флюорометра Qubit (Invitrogen, США). Эквимолярные количества (2 нмоль) каждой библиотеки были объединены и секвенированы с помощью системы Illumina MiSeq (Illumina, США) с использованием набора для одноконцевого чтения 150 п. н.
В результате секвенирования были получены файлы формата FASTQ по каждой клеточной линии. Их биоинформационную обработку проводили на платформе GenXPro omiRas (Германия) [16], в результате чего получены данные в виде таблиц, содержащих нормализованное на 105 прочтений количество микро-РНК наименования.
Поиск генов, экспрессия которых регулируется микро-РНК, также проводили с использованием платформы GenXPro omiRas (Германия) [16]. Для дальнейший работы были отобраны микро-РНК с нормализованной экспрессией (>100 штук на образец). В результате формировался список генов для каждой клеточной линии и экспериментальной точки, экспрессию которых регулируют микро-РНК.
Поиск сигнальных путей, в которые включены данные гены, проводили в базе данных PANTHER (США) (http://www.pantherdb.org/). Так как каждая микро-РНК с равной вероятностью участвует в каждом сигнальном пути, то нормализованное количество транскриптов каждой микро-РНК делили на количество сигнальных путей. Затем суммировали полученные значения по нормализованной экспрессии микро-РНК каждого сигнального пути и получали сумму нормализованных экспрессий (СНЭ) всех микро-РНК, которые участвуют в посттранскрипционной регуляции экспрессии генов, включенных в этот сигнальный путь. Суммы нормализованных экспрессий опыта и контроля сравнивали и представляли в виде log2 FC (двоичный логарифм отношения СНЭ опыта к контролю). На основе полученных данных были построены графики. Для их построения брали те сигнальные пути, значения log2 FC СНЭ которых были больше 0,5 или меньше –0,5 хотя бы в одном из экспериментов (log2 FC>0,5 и log2 FC<–0,5).
Результаты. На рис. 1 для всех трех клеточных линий K562, HL-60, Raji представлено количество клеток с признаками некроза в контроле и после воздействия ионизирующего излучения в дозе 4 Гр. В клеточной линии K562 в контроле число клеток с признаками некроза составляет 7,23%, после однократного облучения в дозе 4 Гр — возрастает до 13,56%. В радиочувствительной клеточной линии HL-60 количество клеток с признаками некроза после облучения в дозе 4 Гр увеличивается в несколько раз по отношению к контролю и составляет 20,5% вместо 5,23%. В клеточной линии Raji доля клеток с признаками некроза после облучения составляет 31,56%.
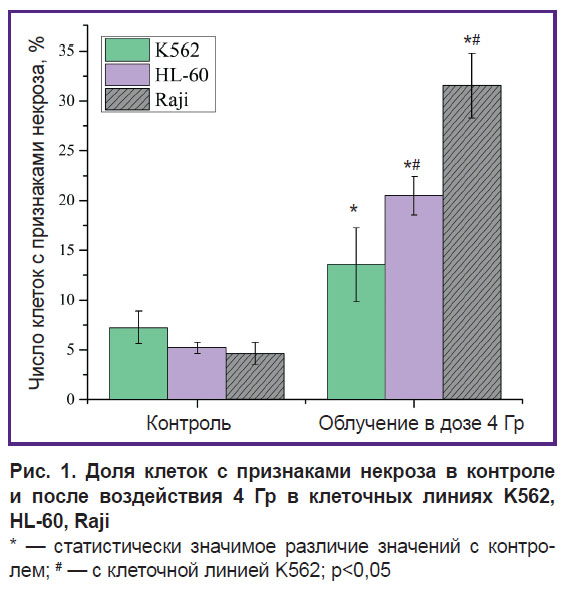
|
Рис. 1. Доля клеток с признаками некроза в контроле и после воздействия 4 Гр в клеточных линиях K562, HL-60, Raji * — статистически значимое различие значений с контролем; # — с клеточной линией K562; p<0,05 |
В результате секвенирования микро-РНК и последующего анализа были выявлены дифференциально-экспрессирующиеся микро-РНК в каждой из линий через 1, 4 и 24 ч после облучения рентгеновским излучением в дозе 4 Гр. Диаграмма Венна (рис. 2) демонстрирует пересечение множеств микро-РНК всех клеточных линий. Через 1 ч после облучения было найдено 4 дифференциально-экспрессирующихся микро-РНК, характерных для всех трех исследуемых линий, — hsa-miR-5701, hsa-miR-590-3p, hsa-miR-5010-3p, hsa-miR-3607-5p (рис. 2, а). Через 4 ч в области пересечения множеств дифференциально-экспрессирующихся микро-РНК оказались только hsa-miR-590-3p, hsa-miR-5010-3p, hsa-miR-3607-5p, а через 24 ч — только одна микро-РНК hsa-miR-590-3p (рис. 2, б, в).
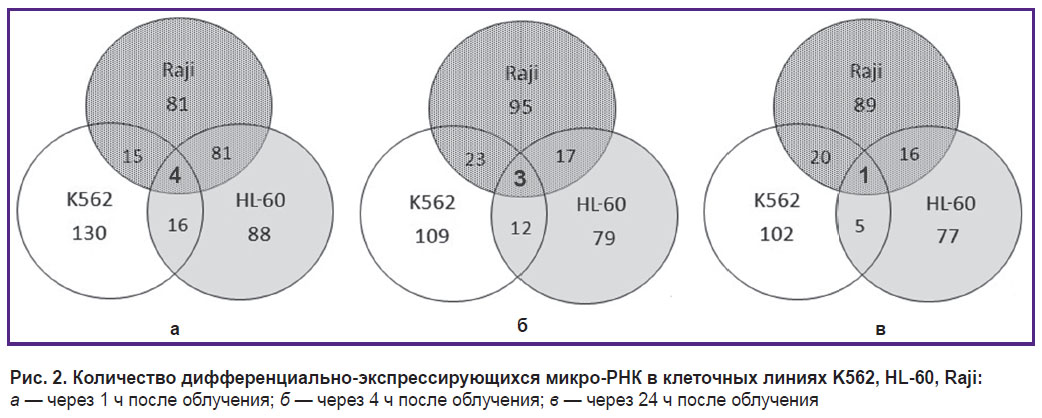
|
Рис. 2. Количество дифференциально-экспрессирующихся микро-РНК в клеточных линиях K562, HL-60, Raji:
а — через 1 ч после облучения; б — через 4 ч после облучения; в — через 24 ч после облучения |
Посттранскрипционная регуляция экспрессии генов посредством микро-РНК обладает рядом особенностей, основной из которых является способность регулировать экспрессию многих генов одной микро-РНК и экспрессию одного гена множеством микро-РНК. Поэтому изучение только дифференциальной экспрессии микро-РНК является недостаточным для понимания различий радиорезистентных и радиочувствительных клеточных линий. Для более глубокого выявления причин радиорезистентности раковых клеток и роли микро-РНК в этом процессе нами был выполнен биоинформационный анализ взаимодействия микро-РНК внутриклеточных сигнальных и метаболических путей (рис. 3). Графики отражают «давление» микро-РНК на функционирование того или иного сигнального пути. Величина давления представлена в виде log2 FC СНЭ. Отрицательные значения log2 FC СНЭ означают, что давление микро-РНК на сигнальный путь возрастает, положительные значения — что давление падает.
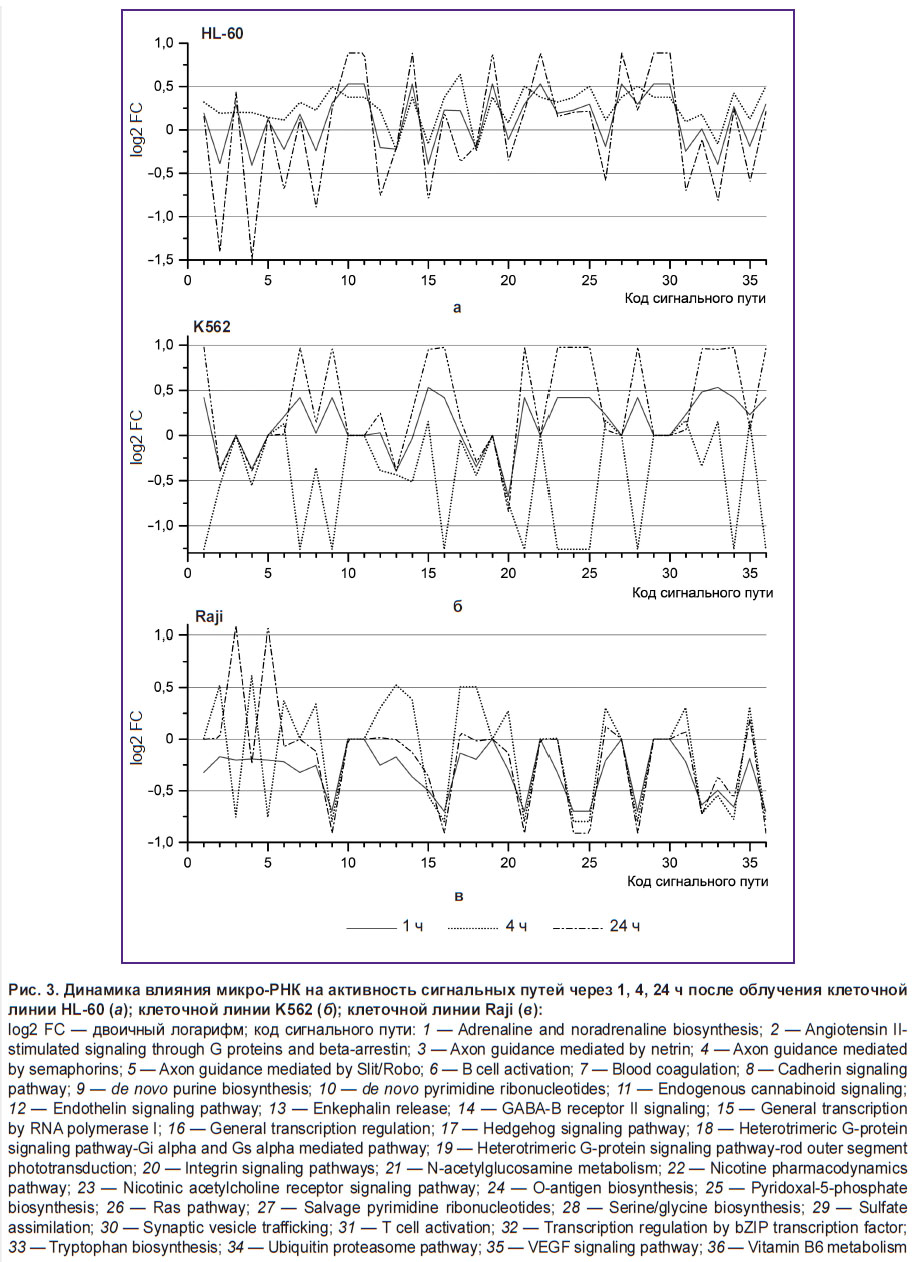
|
Рис. 3. Динамика влияния микро-РНК на активность сигнальных путей через 1, 4, 24 ч после облучения клеточной линии HL-60 (а); клеточной линии K562 (б); клеточной линии Raji (в):
log2 FC — двоичный логарифм; код сигнального пути: 1 — Adrenaline and noradrenaline biosynthesis; 2 — Angiotensin II-stimulated signaling through G proteins and beta-arrestin; 3 — Axon guidance mediated by netrin; 4 — Axon guidance mediated by semaphorins; 5 — Axon guidance mediated by Slit/Robo; 6 — B cell activation; 7 — Blood coagulation; 8 — Cadherin signaling pathway; 9 — de novo purine biosynthesis; 10 — de novo pyrimidine ribonucleotides; 11 — Endogenous cannabinoid signaling; 12 — Endothelin signaling pathway; 13 — Enkephalin release; 14 — GABA-B receptor II signaling; 15 — General transcription by RNA polymerase I; 16 — General transcription regulation; 17 — Hedgehog signaling pathway; 18 — Heterotrimeric G-protein signaling pathway-Gi alpha and Gs |
Из рис. 3 видно, что для клеточной линии К562 характерна нестабильность динамики влияния микро-РНК на активность сигнальных путей. Эта нестабильность проявляется в колебании от отрицательных до положительных значений log2 FC (рис. 3, б). В клеточной линии К562 11 сигнальных путей подвержены таким колебаниям, в клеточной линии Raji — 3 (рис. 3, в), а в клеточной линии HL-60 не обнаружено ни одного такого сигнального пути (рис. 3, а). Наиболее значительные отличия между радиорезистентной клеточной линией К562 и радиочувствительными клеточными линиями HL-60 и Raji наблюдаются в динамике давления микро-РНК на сигнальные пути Integrin signaling pathway (рис. 3, код 20) и General transcription by RNA polymerase I (рис. 3, код 15). Давление микро-РНК на сигнальный путь General transcription by RNA polymerase I в клеточной линии К562 в ходе эксперимента постепенно ослабевает и достигает значения log2 FC=0,9 через 24 ч после облучения (см. рис. 3, б). В клеточных линиях HL-60 и Raji давление микро-РНК на этот сигнальный путь постепенно увеличивается и достигает значений log2 FC=–0,4 … –0,5 через 24 ч после облучения. Давление микро-РНК на Integrin signaling pathway в клеточной линии К562 через 1 ч после облучения снижается (log2 FC=0,51), через 4 ч возвращается к контрольным значениям, а через 24 ч опять снижается и достигает значения log2 FC=1,01 (рис. 3, б). В клеточных линиях HL-60 и Raji давление на этот сигнальный путь увеличено практически на всем протяжении эксперимента и колеблется в районе log2 FC=–0,5 (см. рис. 3, а, в).
Обсуждение. Для эксперимента были выбраны три раковые клеточные линии, отличающиеся радиочувствительностью к гамма-излучению. Самую низкую радиочувствительность демонстрирует клеточная линия К562 — суспензионной хронической миелоидной лейкемии человека, средней радиочувствительностью обладает линия промиелоцитарного лейкоза человека HL-60 и самой высокой — линия лимфобластоидных клеток человека Raji. Смертность клеток линии Raji после однократного облучения гамма-излучением дозой 4 Гр в 2 раза больше, чем смертность клеток линии K562 по сравнению с контролем (31,56 против 13,56% соответственно), а смертность клеток линии HL-60 выше, чем линии К562, на 35% (см. рис. 1). Таким образом, использованные клеточные линии имели сходное происхождение и различную радиочувствительность.
Анализ микро-РНК показал, что микро-РНК hsa-miR-590-3p дифференциально экспрессируется во всех трех клеточных линиях (см. рис. 2), причем в линии К562 значительно слабее, чем в линиях HL-60 и Raji (4–16 случаев на 105 прочтений в К562 против 30–69 случаев на 105 прочтений в HL-60 и Raji). Микро-РНК hsa-miR-590-3p ингибирует митохондриальную дисфункцию и оксидативный стресс в обработанных клетках 1-метил-4-фенил-пиридином линий MES23.5 и SH-SY5Y [17]. Показано, что данная микро-РНК тесно связана со многими нейродегенеративными расстройствами, в том числе играет важную роль при болезни Паркинсона [18–20]. Кроме того, ген JMJD1C идентифицирован в качестве целевого гена hsa-miR-590-3p в организме человека. Подавление экспрессии через hsa-miR-590-3p или нокдаун гена JMJD1C увеличивают экспрессию генов PGC-1-alpha, СРН-1 и TFAM, которые являются ключевыми генами, регулирующими митохондриальную функцию [17]. Ген JMJD1C также играет важную роль в клеточном ответе на повреждение ДНК и необходим для выживания клеток острой миелоидной лейкемии [21, 22]. Радиационное облучение индуцирует увеличение количества hsa-miR-590-3p во всех клеточных линиях, однако в клеточной линии К562 количество такой микро-РНК все равно остается в 2–3 раза меньше, чем в клеточных линиях HL-60 и Raji. Таким образом, динамика и величина экспрессии hsa-miR-590-3p коррелируют с радиочувствительностью раковых клеток.
Анализ СНЭ всех микро-РНК, которые участвуют в посттранскрипционной регуляции экспрессии генов, включенных в отдельные сигнальные пути, позволил выявить различия в давлении микро-РНК на сигнальные пути. В ходе биоинформационного анализа установлены значительные отличия в динамике изменения давления микро-РНК на пути Integrin signaling pathway (см. рис. 3, код 20) и General transcription by RNA polymerase I (см. рис. 3, код 15) между радиорезистентной клеточной линией К562 и радиочувствительными клеточными линиями HL-60 и Raji. Передача сигналов через Integrin signaling pathway тесно связана с развитием, прогрессированием и терапией раковых опухолей [23]. В ряде работ продемонстрирована также взаимосвязь ряда ключевых компонентов Integrin signaling pathway с радиочувствительностью раковых клеток. Так, активация этого пути через адгезию бета-1-интегрина с внеклеточным матриксом способствует увеличению устойчивости к развитию радиационно-индуцированного повреждения генома [24]. Ингибирование передачи сигнала через бета-1-интегрин с использованием антител, наоборот, увеличивает чувствительность раковых клеток к ионизирующей радиации [25]. Наше исследование влияния микро-РНК на сигнальные пути продемонстрировало, что в радиорезистентной клеточной линии К562 ингибирование Integrin signaling pathway после радиационного воздействия ослабевает (см. рис. 3, б, код 15). В клеточных линиях HL-60 и Raji, которые отличаются большей радиочувствительностью, ингибирование Integrin signaling pathway присутствует (см. рис. 3, а, в, код пути 15). Таким образом, анализ влияния микро-РНК на сигнальные пути радиорезистентных и радиочувствительных клеточных линий показал, что в радиорезистентной клеточной линии ослаблено ингибирующее действие микро-РНК на Integrin signaling pathway.
Второй сигнальный путь, найденный нами в результате проделанной работы, на который по-разному влияют микро-РНК в радиорезистентных и радиочувствительных клеточных линиях, это — General transcription by RNA polymerase I. РНК-полимераза I в эукариотических клетках ответственна за синтез рибосомальных РНК (кроме 5S рРНК). Регуляция синтеза рибосомальных РНК связана с синтезом белка. Ингибирование РНК-полимеразы I приводит к активации р53-зависимого апоптоза [26]. В наших экспериментах мы использовали радиорезистентную клеточную линию К562 с нокаутированным геном ТР53. Запуск р53-индуцированного апоптоза в этой клеточной линии невозможен. Генотип клеточной линии К562 позволяет, чтобы этот сигнальный путь находился под сильным давлением микро-РНК без негативных последствий для клетки по сравнению с более радиочувствительными клеточными линиями HL-60 и Raji, у которых есть ген ТР53. Отсутствие гена ТР53 в клеточной линии К562 позволяет этой линии более эффективно, по нашему мнению, перераспределять энергию в пользу репарационных процессов.
Заключение. Радиорезистентные и радиочувствительные клеточные линии имеют отличие в регуляции сигнальных путей посредством микро-РНК. Динамика и величина экспрессии микро-РНК hsa-miR-590-3p коррелируют с радиочувствительностью раковых клеток. Анализ влияния микро-РНК на сигнальные пути радиорезистентных и радиочувствительных клеточных линий показал, что в радиорезистентной клеточной линии К562 ослаблено ингибирующее действие микро-РНК на сигнальные пути Integrin signaling pathway и усилено — на General transcription by RNA polymerase I.
Финансирование исследования. Работа выполнена при поддержке Министерства образования и науки РФ в рамках государственного задания 6.1617.2014/K, программы «УМНИК» и РФФИ №16-04-00504.
Конфликт интересов. У авторов нет конфликта интересов.
Литература
- Guo L., Xiao Y., Fan M., Li J., Wang Y. Profiling global kinome signatures of the radioresistant MCF-7/C6 breast cancer cells using MRM-based targeted proteomics. J Proteome Res 2015; 14(1): 193–201, https://doi.org/10.1021/pr500919w.
- Kim W.-Y., Oh S.H., Woo J.-K., Hong W.K., Lee H.-Y. Targeting heat shock protein 90 overrides the resistance of lung cancer cells by blocking radiation-induced stabilization of hypoxia-inducible factor-1alpha. Cancer Res 2009; 69(4): 1624–1632, https://doi.org/10.1158/0008-5472.can-08-0505.
- Tang Y., Cui Y., Li Z., Jiao Z., Zhang Y., He Y., Chen G., Zhou Q., Wang W., Zhou X., Luo J., Zhang S. Radiation-induced miR-208a increases the proliferation and radioresistance by targeting p21 in human lung cancer cells. J Exp Clin Cancer Res 2016; 35: 7, https://doi.org/10.1186/s13046-016-0285-3.
- Li J.Y., Li Y.Y., Jin W., Yang Q., Shao Z.M., Tian X.S. ABT-737 reverses the acquired radioresistance of breast cancer cells by targeting Bcl-2 and Bcl-xL. J Exp Clin Cancer Res 2012; 31: 102, https://doi.org/10.1186/1756-9966-31-102.
- Filipowicz W., Bhattacharyya S.N., Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat Rev Genet 2008; 9(2): 102–114, https://doi.org/10.1038/nrg2290.
- Zheng L., Zhang Y., Liu Y., Zhou M., Lu Y., Yuan L., Zhang C., Hong M., Wang S., Li X. MiR-106b induces cell radioresistance via the PTEN/PI3K/AKT pathways and p21 in colorectal cancer. J Transl Med 2015; 13: 252, https://doi.org/10.1186/s12967-015-0592-z.
- Ma R., Jiang T., Kang X. Circulating microRNAs in cancer: origin, function and application. J Exp Clin Cancer Res 2012; 31: 38–47, https://doi.org/10.1186/1756-9966-31-38.
- Suzuki H., Maruyama R., Yamamoto E., Kai M. Epigenetic alteration and microRNA dysregulation in cancer. Front Genet 2013; 4: 258, https://doi.org/10.3389/fgene.2013.00258.
- Drakaki A., Iliopoulos D. MicroRNA-gene signaling pathways in pancreatic cancer. Biomed J 2013; 36(5): 200–208, https://doi.org/10.4103/2319-4170.119690.
- Metheetrairut C., Slack F.J. MicroRNAs in the ionizing radiation response and in radiotherapy. Curr Opin Genet Dev 2013; 23(1): 12–19, https://doi.org/10.1016/j.gde.2013.01.002.
- Li G., Qiu Y., Su Z., Ren S., Liu C., Tian Y., Liu Y. Genome-wide analyses of radioresistance-associated miRNA expression profile in nasopharyngeal carcinoma using next generation deep sequencing. PLoS One 2013; 8(12): e84486, https://doi.org/10.1371/journal.pone.0084486.
- Zhang B., Chen J., Ren Z., Chen Y., Li J., Miao X., Song Y., Zhao T., Li Y., Shi Y., Ren D., Liu J. A specific miRNA signature promotes radioresistance of human cervical cancer cells. Cancer Cell Int 2013; 13(1): 118, https://doi.org/10.1186/1475-2867-13-118.
- Su H., Jin X., Zhang X., Xue S., Deng X., Shen L., Fang Y., Xie C. Identification of microRNAs involved in the radioresistance of esophageal cancer cells. Cell Biology Int 2014; 38(3): 318–325, https://doi.org/10.1002/cbin.10202.
- Qu C., Liang Z., Huang J., Zhao R., Su C., Wang S., Wang X., Zhang R., Lee M.H., Yang H. MiR-205 determines the radioresistance of human nasopharyngeal carcinoma by directly targeting PTEN. Cell Cycle 2012; 11(4): 785–796, https://doi.org/10.4161/cc.11.4.19228.
- Mascotti K., McCullough J., Burger S.R. HPC viability measurement: trypan blue versus acridine orange and propidium iodide. Transfusion 2000; 40(6): 693–696, https://doi.org/10.1046/j.1537-2995.2000.40060693.x.
- Müller S., Rycak L., Winter P., Kahl G., Koch I., Rotter B. omiRas: a Web server for differential expression analysis of miRNAs derived from small RNA-Seq data. Bioinformatics 2013; 29(20): 2651–2652, https://doi.org/10.1093/bioinformatics/btt457.
- Wang J., Le T., Wei R., Jiao Y. Knockdown of JMJD1C, a target gene of hsa-miR-590-3p, inhibits mitochondrial dysfunction and oxidative stress in MPP+-treated MES23.5 and SH-SY5Y cells. Cell Mol Biol 2016; 62(3): 39–45.
- Villa C., Fenoglio C., De Riz M., Clerici F., Marcone A., Benussi L., Ghidoni R., Gallone S., Cortini F., Serpente M., Cantoni C., Fumagalli G., Martinelli Boneschi F., Cappa S., Binetti G., Franceschi M., Rainero I., Giordana M.T., Mariani C., Bresolin N., Scarpini E., Galimberti D. Role of hnRNP-A1 and miR-590-3p in neuronal death: genetics and expression analysis in patients with Alzheimer disease and frontotemporal lobar degeneration. Rejuvenation Res 2011; 14(3): 275–281, https://doi.org/10.1089/rej.2010.1123.
- Vaishnavi V., Manikandan M., Tiwary B.K., Munirajan A.K. Insights on the functional impact of microRNAs present in autism-associated copy number variants. PLoS One 2013; 8(2): e56781, https://doi.org/10.1371/journal.pone.0056781.
- Spencer P., Fry R.C., Kisby G.E. Unraveling 50-year-old clues linking neurodegeneration and cancer to cycad toxins: are microRNAs common mediators? Front Genet 2012; 3: 192, https://doi.org/10.3389/fgene.2012.00192.
- Chen M., Zhu N., Liu X., Laurent B., Tang Z., Eng R., Shi Y., Armstrong S.A., Roeder R.G. JMJD1C is required for the survival of acute myeloid leukemia by functioning as a coactivator for key transcription factors. Genes Dev 2015; 29(20): 2123–2139, https://doi.org/10.1101/gad.267278.115.
- Zhu N., Chen M., Eng R., DeJong J., Sinha A.U., Rahnamay N.F., Koche R., Al-Shahrour F., Minehart J.C., Chen C.W., Deshpande A.J., Xu H., Chu S.H., Ebert B.L., Roeder R.G., Armstrong S.A. MLL-AF9- and HOXA9-mediated acute myeloid leukemia stem cell self-renewal requires JMJD1C. J Clin Invest 2016; 126(3): 997–1011, https://doi.org/10.1172/jci82978.
- Desgrosellier J.S., Cheresh D.A. Integrins in cancer: biological implications and therapeutic opportunities. Nat Rev Cancer 2010; 10(1): 9–22, https://doi.org/10.1038/nrc2748.
- Cordes N., Seidler J., Durzok R., Geinitz H., Brakebusch C. β1-integrin-mediated signaling essentially contributes to cell survival after radiation-induced genotoxic injury. Oncogene 2006; 25(9): 1378–1390, https://doi.org/10.1038/sj.onc.1209164.
- Eke I., Deuse Y., Hehlgans S., Gurtner K., Krause M., Baumann M., Shevchenko A., Sandfort V., Cordes N. β1 integrin/FAK/cortactin signaling is essential for human head and neck cancer resistance to radiotherapy. J Clin Invest 2012; 122(4): 1529–1540, https://doi.org/10.1172/jci61350.
- Bywater M.J., Poortinga G., Sanij E., Hein N., Peck A., Cullinane C., Wall M., Cluse L., Drygin D., Anderes K., Huser N., Proffitt C., Bliesath J., Haddach M., Schwaebe M.K., Ryckman D.M., Rice W.G., Schmitt C., Lowe S.W., Johnstone R.W., Pearson R.B., McArthur G.A., Hannan R.D. Inhibition of RNA polymerase I as a therapeutic strategy to promote cancer-specific activation of p53. Cancer Cell 2012; 22(1): 51–65, https://doi.org/10.1016/j.ccr.2012.05.019.








