Транзиторное повышение проницаемости гематоэнцефалического барьера путем интракаротидного введения озонированного физиологического раствора
Цель исследования — оценить воздействие озонированного физиологического раствора с различной концентрацией в нем озона на динамику проницаемости гематоэнцефалического барьера (ГЭБ) в эксперименте.
Материалы и методы. Исследование ультраструктуры сенсомоторной зоны коры головного мозга животных для оценки состояния ГЭБ проведено на половозрелых аутбредных крысах линии Wistar (n=93). В контрольной серии интракаротидно вводился 0,9% физиологический раствор, во опытных сериях — озонированный физиологический раствор (ОФР) с концентрацией озона 0,7 и 3,5 мг/л. Морфологически исследовали ткань сенсомоторной коры головного мозга крыс через 15 мин, 1, 14 и 30 сут после введения растворов. Анализ проницаемости ГЭБ оценивали по состоянию астроглии, базальной мембраны и межклеточных контактов эндотелия капилляров.
Результаты. Установлено, что после однократного введения ОФР с концентрацией озона 0,7 мг/л через 15 мин происходило транзиторное нарушение проницаемости ГЭБ, что проявлялось частичной редукцией межэндотелиальных плотных контактов (tightjunction), расширением межклеточных щелей, набуханием базальной мембраны и части отростков астроцитов в перикапиллярной области. Изменения ультраструктуры нейронов носили адаптивный характер в ответ на воздействие окислительного фактора и характеризовались увеличением активности белоксинтезирующих органелл. Через 1 сут после воздействия ОФР нарастало повышение проницаемости ГЭБ, на 14-е сутки структура ГЭБ частично нормализовалась (эндотелий капилляров имел плотные межклеточные контакты), а к 30-м суткам ультраструктура ГЭБ и микроциркуляция восстанавливались. Увеличение концентрации озона до 3,5 мг/л в ОФР, вводимом интракаротидно, также приводило к повышению проницаемости ГЭБ, однако его восстановления к 30-м суткам не наблюдалось. Кроме того, изменения сосудов микроциркуляторного русла, ультраструктуры нейронов и глии были более выражены.
Заключение. Однократное интракаротидное введение ОФР с концентрацией озона 0,7 мг/л вызывает увеличение проницаемости ГЭБ на период 1 сут, что сможет обеспечить более эффективное воздействие препаратов на клетки головного мозга во все фазы клеточного цикла.
Основными условиями проникновения химиопрепаратов через неповрежденный гематоэнцефалический барьер (ГЭБ) являются малый размер молекул вещества, степень его ионизации и растворимость в жирах [1, 2]. Ряд химиопрепаратов не отвечают как минимум одному из этих требований, и поэтому их эффективность при лечении злокачественных опухолей головного мозга невысокая. Наличие опухоли головного мозга, по данным ряда авторов, вызывает повышение проницаемости церебральных сосудов различной степени в пределах новообразований,однако полного нарушения ГЭБ не наблюдается [3].
Реальным путем достижения препаратами клеток может быть увеличение проницаемости для них ГЭБ. В литературе описаны методы, способные вызвать временное раскрытие ГЭБ: гипертермия, повышение давления в мозговых сосудах выше 90 мм рт. ст., вдыхание газовой смеси с повышенным содержанием двуокиси углерода, введение гистамина, блокаторов кальциевых каналов, лейкотриена С4, брадикинина, интракаротидное введение гиперосмолярных растворов маннитола, изучаются физиологические транспортные механизмы [4–8]. Но большинство из них имеют ряд недостатков, среди которых наличие серьезных осложнений и короткое терапевтическое окно — время, на протяжении которого облегчается доставка препаратов к опухоли через нарушенный ГЭБ [1]. Поэтому для повышения эффективности действия препаратов остается актуальной разработка новых эффективных методов, позволяющих длительно, но обратимо изменять проницаемость ГЭБ.
Известно, что свободнорадикальные соединения способствуют нарушению ГЭБ. Исследована возможность использования озона в качестве источника активных форм кислорода для повышения проницаемости ГЭБ [9, 10].
Цель исследования — оценить воздействие озонированного физиологического раствора с различной концентрацией в нем озона на динамику проницаемости гематоэнцефалического барьера в эксперименте.
Материалы и методы. Исследование проведено на половозрелых аутбредных крысах линии Wistar (n=93) массой тела 320±10 г с соблюдением правил обращения с животными согласно Приказу МЗ РФ №708н от 23.08.2010 «Об утверждении правил лабораторной практики» и в соответствии с этическими принципами Европейской конвенции по защите позвоночных животных, используемых для экспериментальных и других научных целей (принятой в Страсбурге 18.03.1986 г. и подтвержденной в Страсбурге 15.06.2006 г.). Получено разрешение Этического комитета Приволжского федерального медицинского исследовательского центра на работу. Выполнено три серии экспериментов, в которых изучалась ультраструктура сенсомоторной зоны коры головного мозга животных. В I (контрольной) серии интракаротидно вводился 0,9% физиологический раствор (ФР), во II серии — озонированный физиологический раствор (ОФР) с концентрацией озона 0,7 мг/л, в III серии — ОФР с концентрацией озона 3,5 мг/л.
Барботаж ФР озоном длительностью 40 мин осуществлялся прибором «Квазар» (КБ «Квазар», Россия). Концентрацию растворенного озона в ФР определяли с помощью анализатора озона в жидких средах ИКОЖ-5 (Россия). ОФР через катетер вводили в общую сонную артерию со скоростью 1 мл/мин в объеме 1 мл на 300 г массы животного под внутрибрюшинным нембуталовым наркозом (35 мг/кг). После инфузии растворов сонная артерия перевязывалась. Эвтаназию осуществляли одномоментной декапитацией животного в условиях нембуталового наркоза (45 мг/г).
Для морфологических исследований иссекали ткань сенсомоторной коры головного мозга крыс (по атласу G. Paxinos и C. Watson [11]) на стороне введения ОФР либо ФР. Исследование проводили через 15 мин, 1, 14 и 30 сут после введения ОФР. Методом оценки состояния ГЭБ при введении растворов выбрана трансмиссионная электронная микроскопия. Подготовку проб для электронной микроскопии выполняли по стандартной методике: фиксация 2,5% раствором глютарового альдегида и 1% раствором осмиевой кислоты. Заливку осуществляли в смеси эпоксидных смол аралдита и эпона 812. Ультратонкие срезы готовили на ультратоме Ultracut (Reichert-Jung, Австрия) и просматривали на трансмиссионном электронном микроскопе Morgagni 268D (FEI, США). Морфометрию выполняли с помощью программы АnalySIS.
Проницаемость ГЭБ оценивали по состоянию астроглии, базальной мембраны и межклеточных контактов эндотелия капилляров, поскольку известно, что в церебральном эндотелии, формирующем ГЭБ, ключевым морфофункциональным элементом, определяющим барьерные, транспортные и сигнальные функции, являются межклеточные плотные контакты. Их мобильность, изменение архитектурной организации позволяют в короткие сроки менять проницаемость ГЭБ [12, 13].
Статистическую обработку проводили с помощью программы Statistica 6.0 с использованием критерия Манна–Уитни для независимых выборок. Достоверными считали различия при уровне значимости р<0,05.
Результаты. Электронно-микроскопическое исследование ультраструктуры неокортекса при интракаротидном введении ФР не выявило изменений проницаемости ГЭБ. Диаметр капилляров составлял 3,24±0,13 мкм (см. таблицу).

|
Диаметр капилляров сенсомоторной зоны неокортекса белых крыс после введения озонированного физиологического раствора, мкм (М±SD) |
После введения ОФР с концентрацией 0,7 мг/л в неокортексе животных через 15 мин отмечалось очаговое набухание эндотелия капилляров и базальной мембраны. В участках набухания обнаружена редукция межэндотелиальных плотных контактов. Выявлено набухание отдельных отростков астроцитов в перикапиллярной области. Капилляры имели более широкий просвет, чем в контроле (диаметр 4,65±0,38 мкм, p<0,001). В просветах сосудов просматривались тонкодисперсный осмиофильный материал и эритроциты. Значительная часть светлых нейронов содержала ядрышко, в некоторых из них обнаружено эктопирование его к кариолемме, расширение цистерн комплекса Гольджи, незначительные инвагинации ядерной мембраны.
Через 1 сут после введения ОФР с концентрацией озона 0,7 мг/л состояние ГЭБ свидетельствовало о повышении его проницаемости: эндотелий значительного числа капилляров был набухший, имел неровные контуры, темную цитоплазму, нечеткие клеточные мембраны (рис. 1, а). В цитоплазме наблюдались небольшие вакуоли и пиноцитозные пузырьки в сублеммальной области. Межэндотелиальные щели были расширены (более 20 нм). Базальная мембрана — набухшая, извитая, местами разволокнена, на значительном протяжении расплавлена. Этим зонам, как правило, подлежат набухшие и увеличенные в размерах отростки прилегающих к капилляру астроцитов. Наряду с этим выявлены астроциты обычного вида без признаков набухания. Ухудшалась и микроциркуляция: большинство сосудов имели суженный просвет (см. таблицу), содержали плазму. В части сосудов, содержащих эритроциты, наблюдалась их адгезия к эндотелию. Светлые нейроны имели четкую ультраструктуру: ядро содержало эухроматин, в половине наблюдений нейроны были с хорошо выраженным ядрышком. В некоторых из них ядрышко эктопировано к кариолемме, что сопровождалось активацией комплекса Гольджи (см. рис. 1, б). В отдельных нейронах наблюдалась инвагинация ядра, в цитоплазме этих нейронов отмечено удлинение и расширение цистерн гранулярного эндоплазматического ретикулума и комплекса Гольджи. Митохондрии были с плотным матриксом, наблюдалось много рибосом и полисом, первичные и вторичные лизосомы. Наряду со светлыми нейронами встречались гиперхромные нейроны.
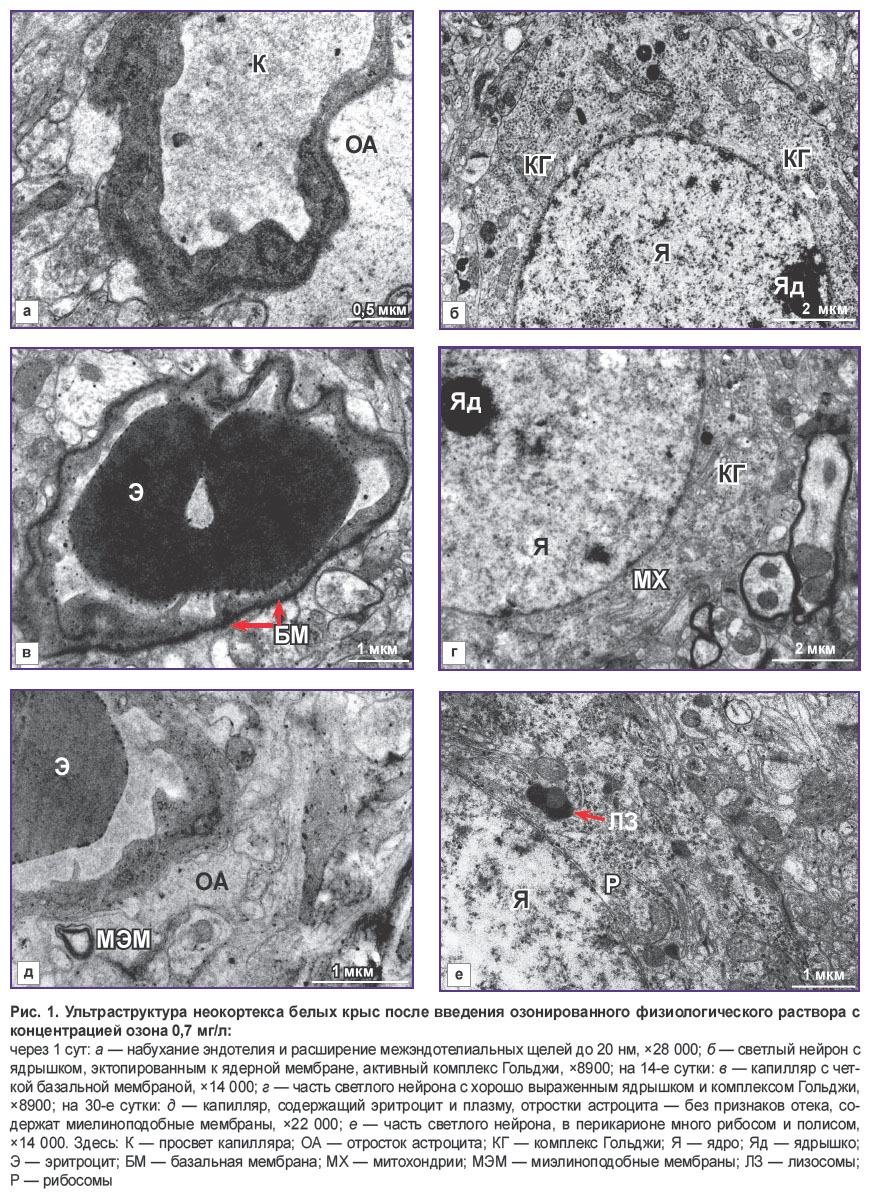
|
Рис. 1. Ультраструктура неокортекса белых крыс после введения озонированного физиологического раствора с концентрацией озона 0,7 мг/л:
через 1 сут: а — набухание эндотелия и расширение межэндотелиальных щелей до 20 нм, ×28 000; б — светлый нейрон с ядрышком, эктопированным к ядерной мембране, активный комплекс Гольджи, ×8900; на 14-е сутки: в — капилляр с четкой базальной мембраной, ×14 000; г — часть светлого нейрона с хорошо выраженным ядрышком и комплексом Гольджи, ×8900; на 30-е сутки: д — капилляр, содержащий эритроцит и плазму, отростки астроцита — без признаков отека, содержат миелиноподобные мембраны, ×22 000; е — часть светлого нейрона, в перикарионе много рибосом и полисом, ×14 000. Здесь: К — просвет капилляра; ОА — отросток астроцита; КГ — комплекс Гольджи; Я — ядро; Яд — ядрышко; Э — эритроцит; БМ — базальная мембрана; МХ — митохондрии; МЭМ — миэлиноподобные мембраны; ЛЗ — лизосомы; Р — рибосомы |
На 14-й день после введения ОФР с концентрацией озона 0,7 мг/л у животных этой серии электронно-микроскопический анализ показал восстановление структуры ГЭБ: эндотелий капилляров в большинстве своем стал обычного вида, имел плотные межклеточные соединения. В некоторых эндотелиоцитах выявлены набухшие митохондрии. Базальная мембрана четко выражена в большей части наблюдений, местами — набухшая (рис. 1, в). Астроглия была в большинстве случаев без признаков набухания. Отмечалось восстановление микроциркуляции (см. таблицу), в просветах капилляров находился тонкодисперсный осмиофильный материал и эритроциты. В отдельных случаях обнаружены фагоцитирующие перициты. Светлые нейроны — обычного вида: ядра содержали эухроматин, ядрышко, комплексы Гольджи (рис. 1, г), иногда набухшие митохондрии. В то же время встречались единичные дегенеративные формы нейронов.
Через 30 сут после введения ОФР с концентрацией озона 0,7 мг/л не выявлено нарушения проницаемости ГЭБ. Периваскулярное пространство не имело признаков набухания. В единичных случаях в ножках астроглии отмечались миелиноподобные мембраны (рис. 1, д). Значительное большинство микрососудов имело широкий просвет и содержало тонкодисперсный осмиофильный материал и эритроциты. Светлые нейроны находились в разной степени активности и изменения ультраструктуры: большинство ядер содержали эухроматин, в цитоплазме наблюдалось много рибосом и полисом, комплекс Гольджи был хорошо выражен, встречались первичные и вторичные лизосомы (рис. 1, е).
Анализ ультраструктуры ГЭБ капилляров сенсомоторной коры головного мозга крыс линии Wistar после введения ОФР с концентрацией озона 3,5 мг/л через 1 сут показал набухание эндотелия капилляров и базальной мембраны (рис. 2). В эндотелиоцитах наблюдалось повышенное содержание пиноцитозных пузырьков, набухшие митохондрии. Межэндотелиальные щели были расширены (около 25 нм).
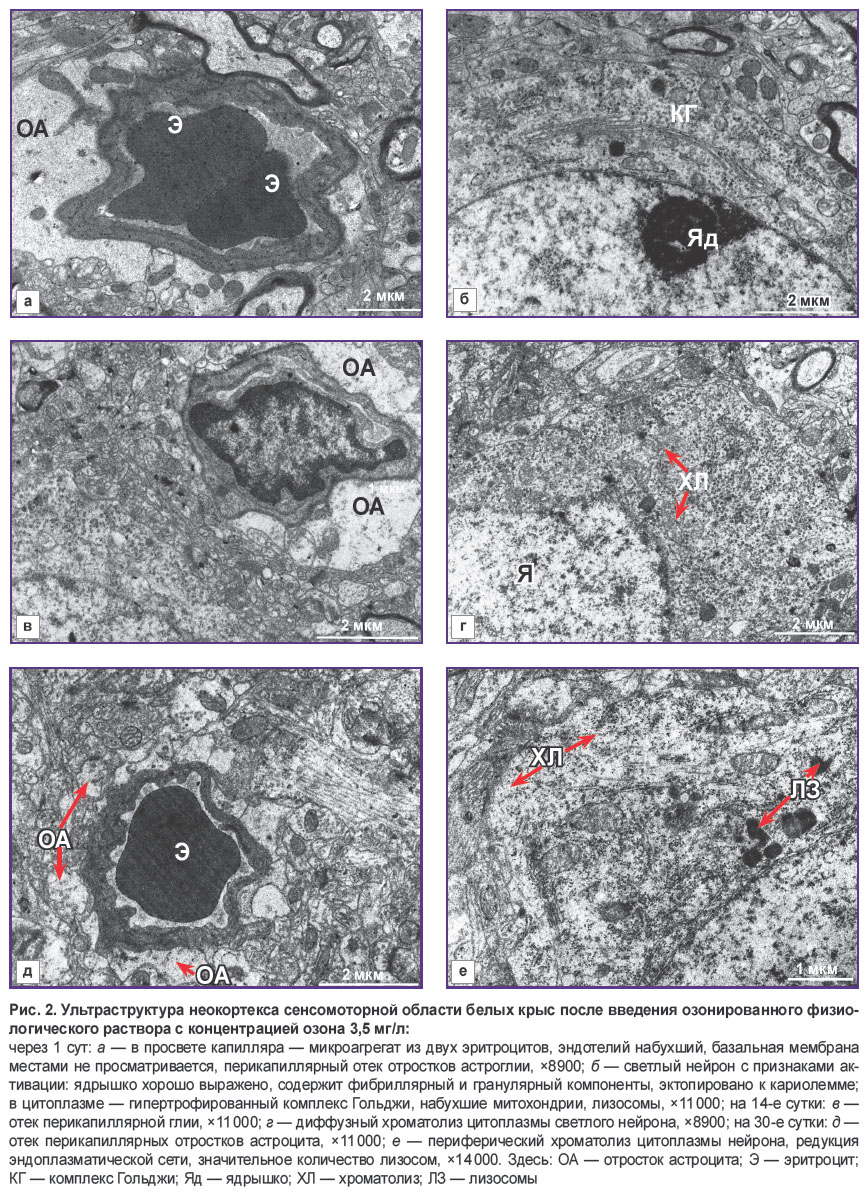
|
Рис. 2. Ультраструктура неокортекса сенсомоторной области белых крыс после введения озонированного физиологического раствора с концентрацией озона 3,5 мг/л:
через 1 сут: а — в просвете капилляра — микроагрегат из двух эритроцитов, эндотелий набухший, базальная мембрана местами не просматривается, перикапиллярный отек отростков астроглии, ×8900; б — светлый нейрон с признаками активации: ядрышко хорошо выражено, содержит фибриллярный и гранулярный компоненты, эктопировано к кариолемме; в цитоплазме — гипертрофированный комплекс Гольджи, набухшие митохондрии, лизосомы, ×11 000; на 14-е сутки: в — отек перикапиллярной глии, ×11 000; г — диффузный хроматолиз цитоплазмы светлого нейрона, ×8900; на 30-е сутки: д — отек перикапиллярных отростков астроцита, ×11 000; е — периферический хроматолиз цитоплазмы нейрона, редукция эндоплазматической сети, значительное количество лизосом, ×14 000. Здесь: ОА — отросток астроцита; Э — эритроцит; КГ — комплекс Гольджи; Яд — ядрышко; ХЛ — хроматолиз; ЛЗ — лизосомы |
Отмечено увеличение периваскулярных отростков астроглии, их набухание и вакуолизация (рис. 2, а). В ряде случаев увеличенные отростки астроцитов, как футляр, окружали капилляр, сдавливая его. У большинства капилляров диаметр был существенно меньше, чем у животных, получавших физиологический раствор (p<0,01). В капиллярах находилась как обычная кровь, так и плазма, микроагрегаты эритроцитов, в единичных наблюдениях выявлена адгезия эритроцитов к эндотелию. Перициты были с признаками фагоцитоза. Часть светлых нейронов была обычного вида, часть имела признаки незначительного диффузного хроматолиза (редукция части органелл, набухание митохондрий, удлинение цистерн гранулярного эндоплазматического ретикулума, уменьшение рибосом и полисом), отмечались нейроны с признаками активации белоксинтезирующих органелл (увеличение ядрышка, эктопирование его к кариолемме, пролиферация и гипертрофия комплекса Гольджи, увеличение рибосом и полисом) (рис. 2, б). Наряду с этим встречались пикнотичные и гиперхромные нейроны.
Через 14 сут у животных после введения ОФР с концентрацией озона 3,5 мг/л нарушение проницаемости ГЭБ сохранялось и проявлялось набуханием эндотелия капилляров, базальной мембраны, лизисом отдельных ее участков и извитым контуром, отеком периваскулярной глии (рис. 2, в). Просвет сосудов оставался более узким, чем в контроле (p<0,01). 1/3 светлых нейронов содержали ядрышки, ядра с инвагинациями, иногда глубокими. Отмечался диффузный хроматолиз отдельных нейронов (рис. 2, г).
На 30-е сутки у этих животных нарушение структуры ГЭБ все еще сохранялось в виде набухания эндотелия, базальной мембраны, периваскулярной астроглии (рис. 2, д), очагового расплавления базальной мембраны, что вызывало изменения ультраструктуры нейронов (рис. 2, е) — появление дегенеративных форм (хроматолиз цитоплазмы, темных нейронов).
Обсуждение. Анализ ультраструктуры ГЭБ проводили, основываясь на данных литературы, согласно которым признаками нарушения проницаемости ГЭБ являются увеличение размеров отростков астроцитов, прилежащих к капиллярам, неровность контура и неравномерная толщина базальной мембраны, увеличенное количество пиноцитозных пузырьков в эндотелии, нарушение межэндотелиальных контактов.
Результаты исследования показали, что после однократного введения ОФР с концентрациейозона 0,7 мг/л через 15 мин происходило нарушение проницаемости ГЭБ, которое проявлялось частичной редукцией межэндотелиальных плотных контактов (tight junction), расширением межклеточных щелей, набуханием базальной мембраны и части отростков астроцитов в перикапиллярной области. Контакты — это динамичные структуры, способные за счет ремоделирования цитоскелета изменять размеры межклеточных щелей в течение минут, а перикапиллярная реакция астроцитов, прилежащих к капиллярам, свидетельствует о повышении проницаемости стенок капилляров [14, 15]. Изменения ультраструктуры нейронов носили адаптивный характер в ответ на воздействие окислительного фактора и характеризовались увеличением активности белоксинтезирующих органелл. Отмечалось улучшение микроциркуляции по сравнению с контрольными животными, о чем свидетельствовало расширение просвета капилляров.
Через 1 сут после воздействия ОФР нарастало повышение проницаемости ГЭБ: расширение межэндотелиальных щелей до 20 нм, выраженное набухание перикапиллярной астроглии. На фоне повышения проницаемости ГЭБ отмечалось ухудшение микроциркуляции. На 14-е сутки структура ГЭБ частично нормализовалась: эндотелий капилляров имел плотные межклеточные контакты. Сохранялось небольшое набухание отростков астроцитов. Отмечалось восстановление микроциркуляции. К 30-м суткам ультраструктура ГЭБ и микроциркуляция восстанавливались.
Повышение концентрации озона в ОФР, вводимом интракаротидно, до дозы 3,5 мг/л также приводило к повышению проницаемости ГЭБ, однако его восстановление не наблюдалось к 30-м суткам. Кроме того, изменения микроциркуляции, ультраструктуры нейронов и глии были более выражены.
Таким образом, интракаротидное введение ОФР в концентрации 0,7 мг/л вызывает длительное (до суток) транзиторное повышение проницаемости ГЭБ с последующим его частичным восстановлением к14-м суткам и полным восстановлением к 30-м суткам. Временное повышение проницаемости ГЭБ имеет перспективы широкого клинического применения как в нейроонкологии при химиотерапии опухолей головного мозга, так и в неврологии в плане направленной доставки лекарственных препаратов, плохо проникающих через интактный ГЭБ в очаги поражения головного мозга.
Заключение. Интракаротидное введение озонированного физиологического раствора с концентрацией озона 0,7 мг/л вызывает длительное (до суток) транзиторное повышение проницаемости ГЭБ, что может обеспечить более эффективное воздействие лекарственных препаратов на клетки головного мозга во все фазы клеточного цикла, улучшить результаты лечения опухолей головного мозга и ряда неврологических заболеваний.
Финансирование исследования. Исследование не финансировалось какими-либо источниками.
Конфликт интересов, связанный с данным исследованием, у авторов отсутствует.
Литература
- Blanchette M., Tremblay L., Lepage M., Fortin D. Impact of drug size on brain tumor and brain parenchyma delivery after a blood–brain barrier disruption. J Cereb Blood Flow Metab 2014; 34(5): 820–826, https://doi.org/10.1038/jcbfm.2014.14.
- Shilo M., Sharon A., Baranes K., Motiei M., Lellouche J.P., Popovtzer R. The effect of nanoparticle size on the probability to cross the blood-brain barrier: an in-vitro endothelial cell model. J Nanobiotechnology 2015; 13: 19, https://doi.org/10.1186/s12951-015-0075-7.
- Шамаев М.И. Изменение мозговых сосудов при глиомах полушарий головного мозга. Автореф. дис. … докт. мед. наук. Киев; 1983.
- Chen Y., Liu L. Modern methods for delivery of drugs across the blood-brain barrier. Adv Drug Deliv Rev 2012; 64(7): 640–665, https://doi.org/10.1016/j.addr.2011.11.010.
- Kiviniemi V., Korhonen V., Kortelainen J., Rytky S., Keinänen T., Tuovinen T., Isokangas M., Sonkajärvi E., Siniluoto T., Nikkinen J., Alahuhta S., Tervonen O., Turpeenniemi-Hujanen T., Myllylä T., Kuittinen O., Voipio J. Real-time monitoring of human blood-brain barrier disruption. PLoS One 2017; 12(3): e0174072, https://doi.org/10.1371/journal.pone.0174072.
- Leuthardt E.C., Duan C., Kim M.J., Campian J.L., Kim A.H., Miller-Thomas M.M., Shimony J.S., Tran D.D. Hyperthermic laser ablation of recurrent glioblastoma leads to temporary disruption of the peritumoral blood brain barrier. PLoS One 2016; 11(2): e0148613, https://doi.org/10.1371/journal.pone.0148613.
- Li C.H., Shyu M.K., Jhan C., Cheng Y.W., Tsai C.H., Liu C.W., Lee C.C., Chen R.M., Kang J.J. Gold nanoparticles increase endothelial paracellular permeability by altering components of endothelial tight junctions, and increase blood-brain barrier permeability in mice. Toxicol Sci 2015; 148(1): 192–203, https://doi.org/10.1093/toxsci/kfv176.
- Patel M.M., Patel B.M. Crossing the blood-brain barrier: recent advances in drug delivery to the brain. CNS Drugs 2017; 31(2): 109–133, https://doi.org/10.1007/s40263-016-0405-9.
- Jung Y.S., Lee S.W., Park J.H., Seo H.B., Choi B.T., Shin H.K. Electroacupuncture preconditioning reduces ROS generation with NOX4 down-regulation and ameliorates blood-brain barrier disruption after ischemic stroke. J Biomed Sci 2016; 23: 32, https://doi.org/10.1186/s12929-016-0249-0.
- Rochfort K.D., Collins L.E., McLoughlin A., Cummins P.M. Tumour necrosis factor-α-mediated disruption of cerebrovascular endothelial barrier integrity in vitro involves the production of proinflammatory interleukin-6. J Neurochem 2016; 136(3): 564–572, https://doi.org/10.1111/jnc.13408.
- Paxinos G., Watson C. The rat brain in stereotaxic coordinates. 4th edition. San Diego: Academic Press; 1998.
- Чехонин В.П., Баклаушев В.П., Юсубалиева Г.М., Волгина Н.Е., Гурина О.И. Фундаментальные и прикладные аспекты изучения гематоэнцефалического барьера. Вестник Российской академии медицинских наук 2012; 8: 66–78.
- Erdő F., Hutka B., Dénes L. Function, aging and dysfunction of blood-brain barrier. Crossing the barrier. Orv Hetil 2016; 157(51): 2019–2027.
- Cabezas R., Avila M., Gonzalez J., El-Bachá R.S., Báez E., García-Segura L.M., Jurado Coronel J.C., Capani F., Cardona-Gomez G.P., Barreto G.E. Astrocytic modulation of blood brain barrier: perspectives on Parkinson’s disease. Front Cell Neurosci 2014; 8: 211, https://doi.org/10.3389/fncel.2014.00211.
- Liu X., Sui B., Sun J. Blood-brain barrier dysfunction induced by silica NPs in vitro and in vivo: involvement of oxidative stress and Rho-kinase/JNK signaling pathways. Biomaterials 2017; 121: 64–82, https://doi.org/10.1016/j.biomaterials.2017.01.006.








