Эффект знакомости при распознавании лиц и слов: данные ЭЭГ-исследования
Способность к распознаванию лиц и слов имеет решающее значение для социальной коммуникации. На существование тесной взаимосвязи между процессами распознавания лиц и слов указывают многочисленные исследования, проведенные с использованием методов ЭЭГ, вызванных потенциалов, функциональной МРТ, а также описанные в литературе клинические данные о нарушениях восприятия лиц, сопровождающихся ухудшением в сфере распознавания слов. Вследствие этого выявление общих механизмов распознавания вербальных и невербальных стимулов является актуальным не только для нормальной физиологии и когнитивной нейронауки, но и для клиники агнозий различной этиологии.
Мы предположили, что на ЭЭГ-паттерны, сопровождающие распознавание слов и лиц, будет влиять как фактор типа стимула (слово или лицо), так и фактор знакомости.
Цель исследования — определить ЭЭГ-паттерны, ответственные за распознавание и узнавание зрительного стимула независимо от его специфики, т.е. единого для вербальных (слова) и невербальных (лица) стимулов.
Материалы и методы. Регистрировали ЭЭГ-данные у 26 добровольцев, которым предъявляли сложные зрительные стимулы — изображения лиц с наложенными на них словами, где знакомые (известные) и незнакомые лица и слова комбинировались в равных частях. Задачей наблюдателей было в одной части эксперимента классифицировать лица как знакомые или незнакомые (внимание только на лица), в другой части — таким же образом классифицировать слова (внимание только на слова).
Результаты. Установлен выраженный эффект знакомости: амплитуда компонента вызванных потенциалов N250 во фронтальных областях мозга была значимо больше в ответах на незнакомые стимулы (как лица, так и слова) по сравнению со знакомыми. Мы также обнаружили влияние инструкции на ответы (внимание на лица или слова): амплитуда компонента N400 была больше в ответах с инструкцией «внимание на слова» по сравнению с инструкцией «внимание на лица»; этот эффект также был более выражен во фронтальных отведениях.
Заключение. На ранних этапах процесса распознавания зрительных стимулов ответы, регистрируемые методом вызванных потенциалов, модулируются знакомостью стимулов (т.е. представленностью в долговременной памяти), а не их типом (лицо/слово). На более поздних этапах обработки выполняется, по-видимому, категоризация стимулов, обусловленная их модальностью (вербальный или невербальный).
Введение
Способность к распознаванию лиц и слов имеет решающее значение для социальной коммуникации. Согласно распространенному в литературе мнению, за распознавание написанных слов отвечают преимущественно области, расположенные в левом полушарии, а за распознавание лиц — в правом [1–4 и др.]. Многочисленные психофизические и нейрофизиологические исследования свидетельствуют, что подобная локализация обусловлена качественными особенностями этих стимулов. Обнаружено, что левое полушарие преимущественно отвечает за распознавание абстрактных категорий, характеризующихся ассоциативными связями между изолированными элементами, а правое полушарие — за распознавание конкретно-предметных категорий, построенных на пространственных отношениях между элементами [5]. Такие данные, казалось бы, позволяют соотнести процесс распознавания лиц с областью, расположенной в фузиформной извилине (fusiform face area) в правом полушарии [6], а процесс обработки слов — с соответствующей областью фузиформной извилины в левом полушарии (visual word form area) [3] и таким образом предположить существование двух независимых высокоуровневых процессов обработки зрительных стимулов.
Однако появляется все больше работ, результаты которых ставят под сомнение такое четкое разделение. Так, например, несмотря на латерализацию функций распознавания слов и лиц, при предъявлении стимулов этого типа обычно активируются оба полушария в ответ на каждый из них и в обоих полушариях есть перекрывающиеся зоны FFA и VWFA [7, 8], следовательно, при распознавании лиц и слов могут быть задействованы одни и те же процессы [9]. На это указывают и приводимые в литературе клинические данные. Например, еще в работах 1990-х гг. были описаны случаи сложных зрительных агнозий, когда нарушения затрагивают как восприятие лиц, так и распознавание объектов и слов [10, 11]. В работе [12] описан случай пациента с агнозией на лица и слова, развившейся после травмы головы в детском возрасте, причем узнавание объектов нарушено не было. К настоящему времени в литературе накопилось довольно много случаев нарушения восприятия лиц в результате травмы головного мозга (так называемая приобретенная прозопагнозия), когда также нарушается восприятие слов (см., например, обзор [13]). Кроме того, получены данные о том, что у пациентов, страдающих алексией, также наблюдается некоторое ухудшение распознавания лиц [14].
Для изучения динамики процессов обнаружения и распознавания лиц широко применяется метод вызванных потенциалов (ВП). На основании ответов мозга на стимулы с изображением лиц, используемые в различных экспериментальных парадигмах, исследователи выделяют несколько этапов обработки. Самый ранний компонент ВП, который в литературе традиционно связывают с перцептивной обработкой изображений лиц, — это так называемый компонент N170, негативная волна с латентностью в диапазоне 130–200 мс, достигающая своего максимума в ответ на стимулы с изображением лиц в затылочно-теменных отведениях [15]. Полагают, что на этапе N170 происходит обнаружение и выделение лица из фона и его первичная обработка, которая не задействует процессы распознавания [16, 17]. Однако в ряде исследований получены свидетельства того, что амплитуда этого компонента модулируется не только типом стимула, но и его представленностью в долговременной памяти, т.е. знакомостью: амплитуда N170 была больше для лиц хорошо известных людей по сравнению с незнакомыми лицами [18–20]. Таким образом, уже на этапе первых 100–200 мс после предъявления стимула с изображением лица, по всей вероятности, запускается процесс его распознавания.
В ряде исследований обнаружено, что предъявляемые визуально слова вызывают более выраженную негативную волну с латентностью около 170 мс (ее называют также компонент N1, или потенциал распознавания — recognition potential) по сравнению с другими стимулами [21–24]. А в работе [25] показано сходство характеристик компонента N170 в ответах на лица и слова.
Следующее временнóе окно, в пределах которого регистрируются ответы, модулируемые знакомостью/незнакомостью стимулов с изображением лиц, — 200–300 мс от начала стимуляции. Амплитуда компонента N250, регистрируемого в этом временнóм диапазоне в экспериментальной парадигме с использованием многократно повторяющихся стимулов (поэтому данный компонент называют также N250r — repetition), больше в ответах на знакомые лица по сравнению с незнакомыми в затылочно-височных (например, P8 [26] или несколько электродов, включая P10 и TP10 [27]), а также в теменных, центральных и лобных отведениях [28, 29]. Этот компонент связывают с процессом активации репрезентации лицевого паттерна для дальнейшего распознавания и рассматривают как своего рода индекс знакомости [30].
Дальнейшая обработка информации о зрительных стимулах и их соотнесение со следами, хранящимися в долговременной памяти, как полагают, происходит примерно спустя 350–450 мс с начала предъявления. Данный процесс связывают с компонентом, регистрируемым в передних областях, пик которого приходится примерно на 400 мс, а амплитуда зависит от того, знаком ли наблюдателю данный стимул или нет, что подтверждается во многих исследованиях, где в качестве стимулов использовались как лица, так и слова [16, 31–34]. В литературе этот компонент называют midfrontal old-new effect [35] или чаще N400f (familiarity), отличая его тем самым от «классического» N400. Последний в большей степени связан с семантической обработкой стимула и контекста, в котором он предъявляется, хотя исследования показывают тесную взаимосвязь этих двух компонентов [36]. Полагают, что компонент N400f является электрофизиологическим маркером обработки информации в семантической памяти: при узнавании знакомого стимула на этом этапе активируется семантическая информация, хранящаяся в долговременной памяти. Результаты работ с использованием лиц и слов в качестве стимулов свидетельствуют о том, что амплитуда и локализация этого компонента могут зависеть от типа стимула, но, тем не менее, он не является модально-специфичным, т.е. регистрируется в одних и тех же экспериментальных условиях для обоих типов стимулов [33, 37]. Следует отметить, что во всех работах, где в качестве стимулов предъявлялись слова и лица, в результате не проводилось непосредственного сопоставления паттернов ответов на оба типа стимулов: либо перед экспериментаторами стояли иные задачи, либо такое сопоставление было невозможно в силу значительного различия между собой вербальных и невербальных стимулов по своим физическим характеристикам. Например, в относительно недавнем исследовании [38] показано, что ВП при припоминании знакомых лиц и слов во многом схожи: разница в паттернах электроэнцефалограммы при припоминании в гораздо большей степени обусловлена статусом стимула в долговременной семантической памяти (знакомый/незнакомый), чем собственно спецификой стимула — лицо это или слово. Однако выводы делаются опосредованно, а интерпретация результатов носит скорее спекулятивный характер: ВП при припоминании знакомых лиц имеют больше общего с ВП при припоминании слов, чем с ВП при припоминании незнакомых лиц.
Цель исследования — определить ЭЭГ-паттерны, ответственные за распознавание и узнавание зрительного стимула независимо от его специфики, т.е. единые для вербальных (слова) и невербальных (лица) стимулов.
Материалы и методы
Для достижения поставленной цели было необходимо разработать комбинированные вербально-невербальные стимулы, обеспечивающие одновременное предъявление лиц и слов, что позволило бы обнаружить общий для любых зрительных стимулов паттерн распознавания. Мы использовали фотографии известных и неизвестных лиц с наложенными на них известными и неизвестными словами. «Неизвестные лица» мужчин и женщин (50/50) были взяты из бесплатных баз: The FEI face database (http://fei.edu.br/~cet/facedatabase.html) и CVL Face Database (http://www.lrv.fri.uni-lj.si/facedb.html). В качестве «известных лиц» выступали изображения знаменитостей (актеров, политиков, спортсменов, музыкантов) с нейтральным выражением лица, взятые из открытого доступа в Интернете. Предварительно отобранные 120 фотографий знаменитостей в черно-белом варианте были предъявлены 40 добровольцам, которые не участвовали в основной части эксперимента. Их задачей было называть, кто изображен на фотографии. В результате были выбраны 80 наиболее узнаваемых фотографий (40 мужчин и 40 женщин).
Все вербальные стимулы представляли собой пятибуквенные слова русского языка. В качестве «известных» выступали слова с частотностью выше 19 употреблений на миллион слов лингвистического корпуса (ipm — instances per million) и средней частотностью 109 ipm (по частотному словарю [39]). «Неизвестные» слова были подобраны на основе интернет-ресурса http://www.zabytye-slova.ru/. Список из 160 слов был представлен шести экспертам (в роли экспертов выступили 6 носителей русского языка с научной степенью по филологии), задачей которых было вычеркнуть из списка знакомые слова. В результате были отобраны сто слов, из которых в свою очередь были вычеркнуты слова, созвучные с названиями международных брендов (например, «кайен»). В итоговый список стимулов вошли 80 неизвестных слов.
На основании отобранных фотографий и слов в программе Photoshop было составлено 160 комбинированных стимулов, которые представляли собой слова, размещенные поверх лиц, и сочетали «знакомые» и «незнакомые» слова и лица в равных долях (рис. 1).

|
Рис. 1. Примеры комбинированных стимулов: лица известных и незнакомых людей с наложенными на них словами — известными и вышедшими из употребления (незнакомыми) |
В исследовании приняли участие 26 добровольцев 19–35 лет, с нормальным или откорректированным до нормального зрением (12 мужчин, 14 женщин). Участники были заранее ознакомлены с экспериментальной процедурой, проинформированы о возможности прекратить эксперимент в любой момент и об анонимности всех полученных данных. Каждый испытуемый подписал письменное согласие на участие в эксперименте. Исследование было проведено в соответствии с Хельсинкской декларацией (2013) и одобрено этическим комитетом Санкт-Петербургского государственного университета.
В ходе исследования участникам поочередно предъявляли 160 черно-белых изображений стимулов на экране компьютера, расположенного на расстоянии 70 см от наблюдателя. Все изображения были выравнены по яркости и размеру (305×408 пикселей) и предъявлялись на сером фоне. Перед каждой пробой в центре экрана появлялась точка фиксации на 800–2300 мс (время представления варьировалось случайным образом), после этого предъявлялся стимул на 200 мс, который сменялся маской белого шума на 2000 мс. Таким образом, длительность одной пробы составляла от 3000 до 4500 мс.
Процедура исследования состояла из двух сессий, в каждой из которых участникам предъявлялось по 80 стимулов. Задачей участников в одной из сессий было отвечать, знакомо им предъявленное слово или нет, не обращая внимания на лицо; в другой сессии — отвечать, знакомо им предъявленное лицо или нет, не обращая внимания на слово. Для ответов использовали джойстик. Стимулы в двух сессиях не повторялись, очередность заданий была сбалансирована между участниками (половина сначала выполняла задание со словами, а затем с лицами; другая половина, наоборот, сначала работала с лицами, а затем со словами). Участники были проинструктированы фиксировать взглядом крестик в центре экрана и постараться не моргать в период от момента предъявления стимула до нажатия на кнопку при ответе.
Для регистрации электроэнцефалограммы и ВП мозга использовали многоканальный электроэнцефалограф «Нейровизор БММ-52» («МКС», Россия). Запись проводили монополярно в 38 отведениях с двумя ушными референтами (А1, А2), с максимальным сопротивлением до 40 кОм. Частота дискретизации сигнала составляла 1000 Гц. Два электрода, регистрирующие движения глаз, располагались у правого глаза. Коррекция глазодвигательных артефактов производилась автоматически в программе «Неокортекс». Помимо электрической активности фиксировалась точность ответов.
Результаты
После первичного анализа показателей ЭЭГ и ответов участников данные трех испытуемых были исключены из анализа по причине большого числа артефактов или, в одном случае, из-за преобладания ответов «незнаком» на стимулы с лицами известных людей. Анализ ВП проводили только для проб с ответами, соответствующими заданным категориям стимулов — «знакомый»/«незнакомый». Все пробы с пропущенным либо неправильным ответом были исключены из дальнейшего анализа. ВП были усреднены отдельно для каждого отведения и для каждого экспериментального условия. На основании литературных данных для анализа амплитуды волны ВП были выбраны три временныʹх окна: 100–200, 200–300 и 350–450 мс. Эти диапазоны соответствуют описанным компонентам N170, N250 (или N250r) и N400 (N400f). В каждом временнóм окне рассчитывали среднее значение амплитуды. В качестве статистического метода использовали двухфакторный дисперсионный анализ с повторными измерениями (RM ANOVA), где в качестве факторов выступали знакомость (знакомый/незнакомый) и тип стимула, относительно которого участнику необходимо было принимать решение (лицо/слово). На рис. 2 представлены усредненные ВП для четырех групп ответов испытуемых на стимулы.
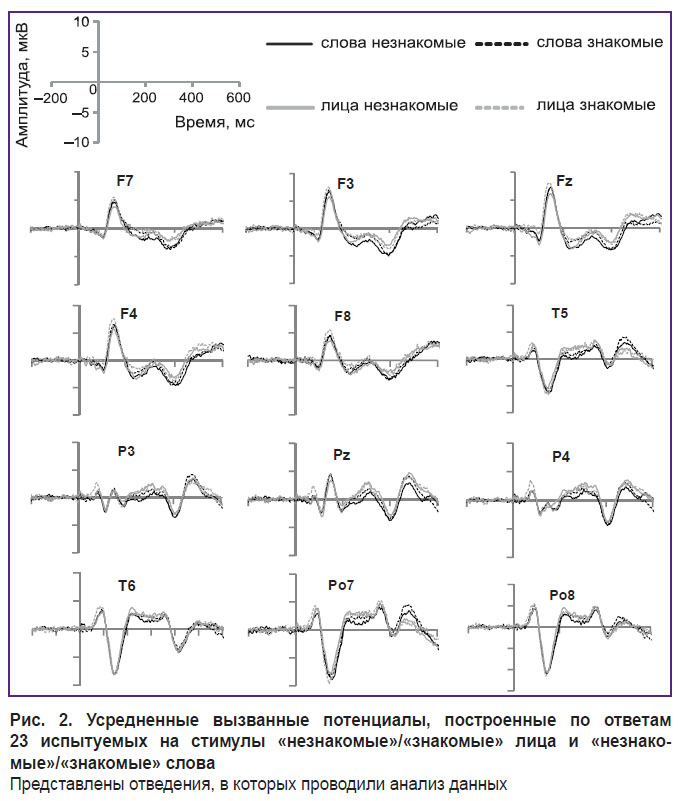
|
Рис. 2. Усредненные вызванные потенциалы, построенные по ответам 23 испытуемых на стимулы «незнакомые»/«знакомые» лица и «незнакомые»/«знакомые» слова
Представлены отведения, в которых проводили анализ данных |
Во всех экспериментальных условиях был обнаружен выраженный компонент N170 (временнóе окно 100–200 мс) в теменно-затылочных отведениях (Pо7, Pо8) и компонент VPP (позитивный вертекс-потенциал) в лобных отведениях (Fz, F3, F4, F7, F8). При этом в теменно-затылочных отведениях не отмечено значимых различий между знакомыми и незнакомыми стимулами (F(1, 22)=1,076; p=0,311), а различия между словами и лицами были на уровне статистической тенденции (F(1, 22)=3,617; p=0,070). В то же время значимым оказалось взаимодействие факторов знакомости и типа стимула (F(1, 22)=4,518; p=0,045). Парные сравнения выявили значимо большую амплитуду компонента N170 для знакомых лиц по сравнению с незнакомыми (F(1, 22)=4,393; p=0,048). Для слов значимых различий между знакомыми и незнакомыми не обнаружено (F(1, 22)=0,766; p=0,391). Во фронтальных отведениях фактор знакомости оказался значим (F(1, 22)=9,297; p=0,006), а тип стимулов — нет (F(1, 22)=0,375; p=0,547). Также было обнаружено значимое взаимодействие факторов знакомости и типа стимула (F(1, 22)=6,010; p=0,023). Парные сравнения выявили значимую разницу в амплитуде компонента VPP для знакомых и незнакомых лиц (F(1, 22)=14,713; p=0,001) — амплитуда компонента была больше для знакомых лиц. Значимых различий в амплитуде VPP между знакомыми и незнакомыми словами не выявлено (F(1, 22)=0,114; p=0,738).
Во временнóм окне 200–300 мс (N250) анализ проводился для фронтальных (Fz, F3, F4, F7, F8), теменных и височных (Pz, P3, P4, T5, T6) отведений. Во фронтальных отведениях обнаружено значимое влияние фактора знакомости (F(1, 22)=29,910; p<0,001) с большей амплитудой волны N250 для незнакомых стимулов; влияния типа стимула не установлено (F(1, 22)=2,674; p=0,116). Также не обнаружено взаимодействия факторов (F(1, 22)=0,486; p=0,493). В теменных отведениях не выявлено влияния знакомости (F(1, 22)=0,940; p=0,343), но обнаружен эффект типа стимула (F(1, 22)=9,360; p=0,006): ответы на незнакомые слова характеризуются большей негативной амплитудой по сравнению с ответами на лица. Взаимодействия факторов не установлено (F(1, 22)=1,881; p=0,184).
Во временном окне 350–450 мс (N400) анализ проводили для фронтальных и теменных отведений. Во фронтальных отведениях не обнаружено эффекта знакомости (F(1, 22)=0,016; p=0,900), но отмечен эффект стимула (F(1, 22)=13,438; p=0,001) — амплитуда компонента N400 была больше в условии «внимание на слова» по сравнению с условием «внимание на лица». Взаимодействия факторов не обнаружено (F(1, 22)=1,910; p=0,181). Для теменных отведений значимых различий не установлено (F<1).
Обсуждение
Результаты проведенного исследования позволили выявить три стадии распознавания испытуемыми зрительных стимулов. ЭЭГ-коррелятом первой стадии является компонент N170 и соответствующий ему по латентному периоду позитивный вертекс-потенциал. На этой стадии, как полагают, происходит первичная перцептивная обработка зрительных стимулов, их структурное декодирование, не задействующее процессы семантической обработки. Обнаруженную в нашем исследовании разницу в опознавании знакомых и незнакомых лиц, выраженную в большей амплитуде компонента N170 для знакомых лиц по сравнению с незнакомыми, можно объяснить тем, что до семантической обработки образ слова, предъявляемый вместе с лицом, неспецифичен и все слова, как знакомые, так и незнакомые, на этом этапе обрабатываются схожим образом; в то время как знакомое лицо уже на этом этапе может модулировать амплитуду ранних компонентов ВП [18–20]. В нашем эксперименте при условии, когда внимание испытуемого было направлено на слова, количество знакомых и незнакомых лиц было одинаковым в каждой группе (знакомые слова/незнакомые слова), поэтому отличия в ответах отсутствуют. При условии, когда внимание было направлено на лица, знакомые и незнакомые лица распределились следующим образом: все знакомые попали в одну группу, а незнакомые — в другую. В результате мы наблюдаем разницу в амплитуде компонента N170.
На следующем этапе, соответствующем временному окну 200–300 мс от начала стимуляции (N250), мы обнаружили достоверное различие в амплитуде ответов на знакомые лица и слова по сравнению с незнакомыми лицами и словами во фронтальных отведениях: знакомые стимулы, вне зависимости от их типа, вызывают более позитивную волну ВП по сравнению с незнакомыми, ответы на которые характеризуются выраженной негативностью. Обычно в исследованиях, направленных на выявление эффекта знакомости, применяется парадигма многократного повторения стимулов («прайминг повторением», repetition priming design), в которой регистрируется компонент N250, его амплитуда в теменных, иногда — центральных и фронтальных отведениях модулируется фактором знакомости/новизны стимула. В большинстве работ отмечается, что знакомые лица при многократном повторении вызывают более негативный ответ по сравнению с незнакомыми, хотя в ряде работ, как и у нас, наблюдается обратный эффект. Например, в исследовании [30] показано, что знакомые (многократно предъявляемые) лица вызывают более позитивную волну по сравнению с незнакомыми в диапазоне 250–450 мс во фронтальных отведениях. В работе [40] также был обнаружен эффект новизны/знакомости во фронтальных отведениях при использовании слов в качестве стимулов. Авторы отмечают, что увеличение амплитуды во фронтальных областях согласуется с данными, полученными при интракраниальной записи активности передних отделов коры, которые свидетельствуют о вкладе префронтальной коры в формирование паттернов ЭЭГ при выполнении задач, задействующих память [41]. Их данные согласуются также с результатами исследований методами функциональной МРТ и позитроннно-эмиссионной томографии, где регистрировалось увеличение активации префронтальной коры в задачах на распознавание лиц [42, 43].
Во временнóм окне 350–450 мс от начала предъявления стимулов (N400) мы получили значимые различия в ответах на стимулы при условии внимания на лица по сравнению с вниманием на слова во фронтальных отведениях. Это может означать, что после активации хранящихся в памяти репрезентаций лиц и слов на следующем этапе обработки зрительной информации происходит ее семантический анализ; в нашем исследовании это отражается в большей амплитуде ответа на слова по сравнению с ответом на лица и обусловлено большей когнитивной нагрузкой в ходе анализа семантики слов. Кроме того, известно [44], что области префронтальной коры участвуют в реализации функции избирательного внимания. В связи с этим полученные нами во фронтальных областях различия между ответами на лица и слова могут также быть результатом влияния инструкции, по которой наблюдателю надо было избирательно направлять внимание либо на слова, игнорируя лица, либо наоборот.
Заключение
С применением метода вызванных потенциалов выявлены ЭЭГ-корреляты последовательных этапов распознавания вербальных (слова) и невербальных (лица) стимулов в экспериментальной парадигме, позволяющей непосредственно соотнести ответы на стимулы этих двух типов и исключающей многократное повторение стимулов. Это позволило оценить влияние фактора знакомости стимулов разных типов (именно как представленности в долговременной памяти) на процесс их распознавания. Можно предположить, что на этапе, соответствующем соотнесению зрительных стимулов с хранящимися в памяти репрезентациями, производится отнесение стимула к одной из категорий — «знакомый»/ «незнакомый» — без учета типа стимула, а на более позднем этапе осуществляется полная семантическая обработка, которая отражается в общем для слов и лиц паттерне N400, однако с большей амплитудой для слов.
Финансирование исследования. Работа поддержана грантом Российского научного фонда, шифр ИАС НИД 37.53.1181.2014.
Конфликт интересов. Авторы подтверждают отсутствие конфликтов интересов, о которых необходимо сообщить.
Литература
- Puce A., Allison T., Asgari M., Gore J.C., McCarthy G. Differential sensitivity of human visual cortex to faces, letter strings, and textures: a functional magnetic resonance imaging study. J Neurosci 1996; 16(16): 5205–5215, https://doi.org/10.1523/jneurosci.16-16-05205.1996.
- McCarthy G., Puce A., Gore J.C., Allison T. Face-specific processing in the human fusiform gyrus. J Cogn Neurosci 1997; 9(5): 605–610, https://doi.org/10.1162/jocn.1997.9.5.605.
- Cohen L., Dehaene S., Naccache L., Lehéricy S., Dehaene-Lambertz G., Hénaff M.A., Michel F. The visual word form area: spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain 2000; 123(2): 291–307, https://doi.org/10.1093/brain/123.2.291.
- Yovel G., Tambini A., Brandman T. The asymmetry of the fusiform face area is a stable individual characteristic that underlies the left-visual-field superiority for faces. Neuropsychologia 2008; 46(13): 3061–3068, https://doi.org/10.1016/j.neuropsychologia.2008.06.017.
- Dien J. A tale of two recognition systems: implications of the fusiform face area and the visual word form area for lateralized object recognition models. Neuropsychologia 2009; 47(1): 1–16, https://doi.org/10.1016/j.neuropsychologia.2008.08.024.
- Kanwisher N., McDermott J., Chun M.M. The fusiform face area: a module in human extrastriate cortex specialized for face perception. J Neurosci 1997; 17(11): 4302–4311, https://doi.org/10.1523/jneurosci.17-11-04302.1997.
- Bouhali F., de Schotten M.T., Pinel P., Poupon C., Mangin J.F., Dehaene S., Cohen L. Anatomical connections of the visual word form area. J Neurosci 2014; 34(46): 15402–15414, https://doi.org/10.1523/jneurosci.4918-13.2014.
- Harris R.J., Rice G.E., Young A.W., Andrews T.J. Distinct but overlapping patterns of response to words and faces in the fusiform gyrus. Cereb Cortex 2015; 26(7): 3161–3168, https://doi.org/10.1093/cercor/bhv147.
- Robinson A.K., Plaut D.C., Behrmann M. Word and face processing engage overlapping distributed networks: evidence from RSVP and EEG investigations. J Exp Psychol Gen 2017; 146(7): 943–961, https://doi.org/10.1037/xge0000302.
- Farah M.J. Cognitive neuropsychology: patterns of co-occurrence among the associative agnosias: implications for visual object representation. Cogn Neuropsychol 1991; 8(1): 1–19, https://doi.org/10.1080/02643299108253364.
- Farah M.J. Dissociable systems for visual recognition: a cognitive neuropsychology approach. In: Kosslyn S.M., Osherson D. (editors). Visual cognition: an invitation to cognitive science. Vol. 2. Cambridge: MIT Press; 1995; p. 101–119.
- Buxbaum L.J., Glosser G., Coslett H.B. Impaired face and word recognition without object agnosia. Neuropsychologia 1998; 37(1): 41–50, https://doi.org/10.1016/s0028-3932(98)00048-7.
- Geskin J., Behrmann M. Congenital prosopagnosia without object agnosia? A literature review. Cogn Neuropsychol 2018; 35(1–2): 4–54, https://doi.org/10.1080/02643294.2017.1392295.
- Behrmann M., Plaut D.C. Bilateral hemispheric processing of words and faces: evidence from word impairments in prosopagnosia and face impairments in pure alexia. Cereb Cortex 2012; 24(4): 1102–1118, https://doi.org/10.1093/cercor/bhs390.
- Rossion B., Jacques C. The N170: understanding the time course of face perception in the human brain. In: Luck S.J., Kappenman A.S. (editors). The Oxford handbook of event-related potential components. Oxford University Press; 2011; p. 15–141, https://doi.org/10.1093/oxfordhb/9780195374148.013.0064.
- Eimer M. Event-related brain potentials distinguish processing stages involved in face perception and recognition. Neurophysiol Clin 2000; 111(4): 694–705, https://doi.org/10.1016/s1388-2457(99)00285-0.
- Engst F.M., Martín-Loeches M., Sommer W. Memory systems for structural and semantic knowledge of faces and buildings. Brain Res 2006; 1124(1): 70–80, https://doi.org/10.1016/j.brainres.2006.09.038.
- Caharel S., Poiroux S., Bernard C., Thibaut F., Lalonde R., Rebai M. ERPs associated with familiarity and degree of familiarity during face recognition. Int J Neurosci 2002; 112(12): 1499–1512, https://doi.org/10.1080/00207450290158368.
- Caharel S., Fiori N., Bernard C., Lalonde R., Rebaï M. The effects of inversion and eye displacements of familiar and unknown faces on early and late-stage ERPs. Int J Psychophysiol 2006; 62(1): 141–151, https://doi.org/10.1016/j.ijpsycho.2006.03.002.
- Wild-Wall N., Dimigen O., Sommer W. Interaction of facial expressions and familiarity: ERP evidence. Biol Psychol 2008; 77(2): 138–149, https://doi.org/10.1016/j.biopsycho.2007.10.001.
- Bentin S., Mouchetant-Rostaing Y., Giard M.H., Echallier J.F., Pernier J. ERP manifestations of processing printed words at different psycholinguistic levels: time course and scalp distribution. J Cogn Neurosci 1999; 11(3): 235–260, https://doi.org/10.1162/089892999563373.
- Maurer U., Brandeis D., McCandliss B.D. Fast, visual specialization for reading in English revealed by the topography of the N170 ERP response. Behav Brain Funct 2005; 1(1): 13, https://doi.org/10.1186/1744-9081-1-13.
- Cao X., Jiang B., Gaspar C., Li C. The overlap of neural selectivity between faces and words: evidences from the N170 adaptation effect. Exp Brain Res 2014; 232(9): 3015–3021, https://doi.org/10.1007/s00221-014-3986-x.
- Zhao J., Li S., Lin S.E., Cao X.H., He S., Weng X.C. Selectivity of N170 in the left hemisphere as an electrophysiological marker for expertise in reading Chinese. Neurosci Bull 2012; 28(5): 577–584, https://doi.org/10.1007/s12264-012-1274-y.
- Rossion B., Joyce C.A., Cottrell G.W., Tarr M.J. Early lateralization and orientation tuning for face, word, and object processing in the visual cortex. Neuroimage 2003; 20(3): 1609–1624, https://doi.org/10.1016/j.neuroimage.2003.07.010.
- Leleu A., Caharel S., Carré J., Montalan B., Snoussi M., Vom Hofe A., Charvin H., Lalonde R., Rebaï M. Perceptual interactions between visual processing of facial familiarity and emotional expression: an event-related potentials study during task-switching. Neurosci Lett 2010; 482(2): 106–111, https://doi.org/10.1016/j.neulet.2010.07.008.
- Tanaka J.W., Curran T., Porterfield A.L., Collins D. Activation of preexisting and acquired face representations: the N250 event-related potential as an index of face familiarity. J Cogn Neurosci 2006; 18(9): 1488–1497, https://doi.org/10.1162/jocn.2006.18.9.1488.
- Begleiter H., Porjesz B., Wang W. Event-related brain potentials differentiate priming and recognition to familiar and unfamiliar faces. Electroencephalogr Clin Neurophysiol 1995; 94(1): 41–49, https://doi.org/10.1016/0013-4694(94)00240-L.
- Miyakoshi M., Nomura M., Ohira H. An ERP study on self-relevant object recognition. Brain Cogn 2007; 63(2): 182–189, https://doi.org/10.1016/j.bandc.2006.12.001.
- Itier R.J., Taylor M.J. Inversion and contrast polarity reversal affect both encoding and recognition processes of unfamiliar faces: a repetition study using ERPs. Neuroimage 2002; 15(2): 353–372, https://doi.org/10.1006/nimg.2001.0982.
- Bentin S., Deouell L.Y. Structural encoding and identification in face processing: ERP evidence for separate mechanisms. Cogn Neuropsychol 2000; 7(1–3): 35–55, https://doi.org/10.1080/026432900380472.
- Yick Y.Y., Wilding E.L. Material-specific neural correlates of memory retrieval. Neuroreport 2008; 19(15): 1463–1467, https://doi.org/10.1097/wnr.0b013e32830ef76f.
- MacKenzie G., Donaldson D.I. Examining the neural basis of episodic memory: ERP evidence that faces are recollected differently from names. Neuropsychologia 2009; 47(13): 2756–2765, https://doi.org/10.1016/j.neuropsychologia.2009.05.025.
- Joyce C.A., Kutas M. Event-related potential correlates of long-term memory for briefly presented faces. J Cogn Neurosci 2005; 17(5): 757–767, https://doi.org/10.1162/0898929053747603.
- Rugg M.D., Allan K. Event-related potential studies of memory. In: Tulving E., Craik F.I.M. (editors). The Oxford handbook of memory. Oxford University Press; 2000; p. 521–537.
- Meyer P., Mecklinger A., Friederici A.D. Bridging the gap between the semantic N400 and the early old/new memory effect. Neuroreport 2007; 18(10): 1009–1013, https://doi.org/10.1097/wnr.0b013e32815277eb.
- Kutas M., Federmeier K.D. Thirty years and counting: finding meaning in the N400 component of the event-related brain potential (ERP). Annu Rev Psychol 2011; 62(1): 621–647, https://doi.org/10.1146/annurev.psych.093008.131123.
- Nie A., Griffin M., Keinath A., Walsh M., Dittmann A., Reder L. ERP profiles for face and word recognition are based on their status in semantic memory not their stimulus category. Brain Res 2014; 1557: 66–73, https://doi.org/10.1016/j.brainres.2014.02.010.
- Ляшевская О.Н., Шаров С.А. Частотный словарь современного русского языка (на материалах Национального корпуса русского языка). M: Азбуковник; 2009.
- Wilding E.L. In what way does the parietal ERP old/new effect index recollection? Int J Psychophysiol 2000; 35(1): 81–87, https://doi.org/10.1016/s0167-8760(99)00095-1.
- Guillem F., N’Kaoua B., Rougier A., Claverie B. Intracranial topography of event-related potentials (N400/P600) elicited during a continuous recognition memory task. Psychophysiology 1995; 32(4): 382–392, https://doi.org/10.1111/j.1469-8986.1995.tb01221.x.
- Courtney S.M., Ungerleider L.G., Keil K., Haxby J.V. Transient and sustained activity in a distributed neural system for human working memory. Nature 1997; 386(6625): 608–611, https://doi.org/10.1038/386608a0.
- Haxby J.V., Ungerleider L.G., Horwitz B., Maisog J.M., Rapoport S.I., Grady C.L. Face encoding and recognition in the human brain. Proc Natl Acad Sci U S A 1996; 93(2): 922–927, https://doi.org/10.1073/pnas.93.2.922.
- Banich M.T., Milham M.P., Atchley R.A., Cohen N.J., Webb A., Wszalek T., Kramer A.F., Liang Z., Barad V., Gullett D., Shah C., Brown C. Prefrontal regions play a predominant role in imposing an attentional ‘set’: evidence from fMRI. Brain Res Cogn Brain Res 2000; 10(1–2): 1–9, https://doi.org/10.1016/s0926-6410(00)00015-x.








